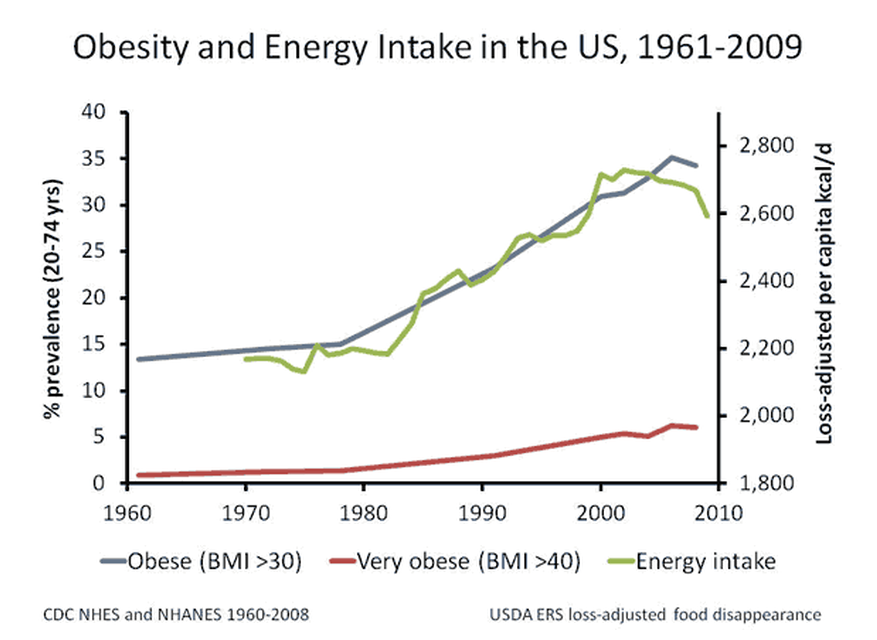
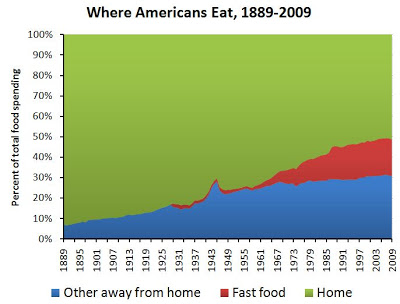
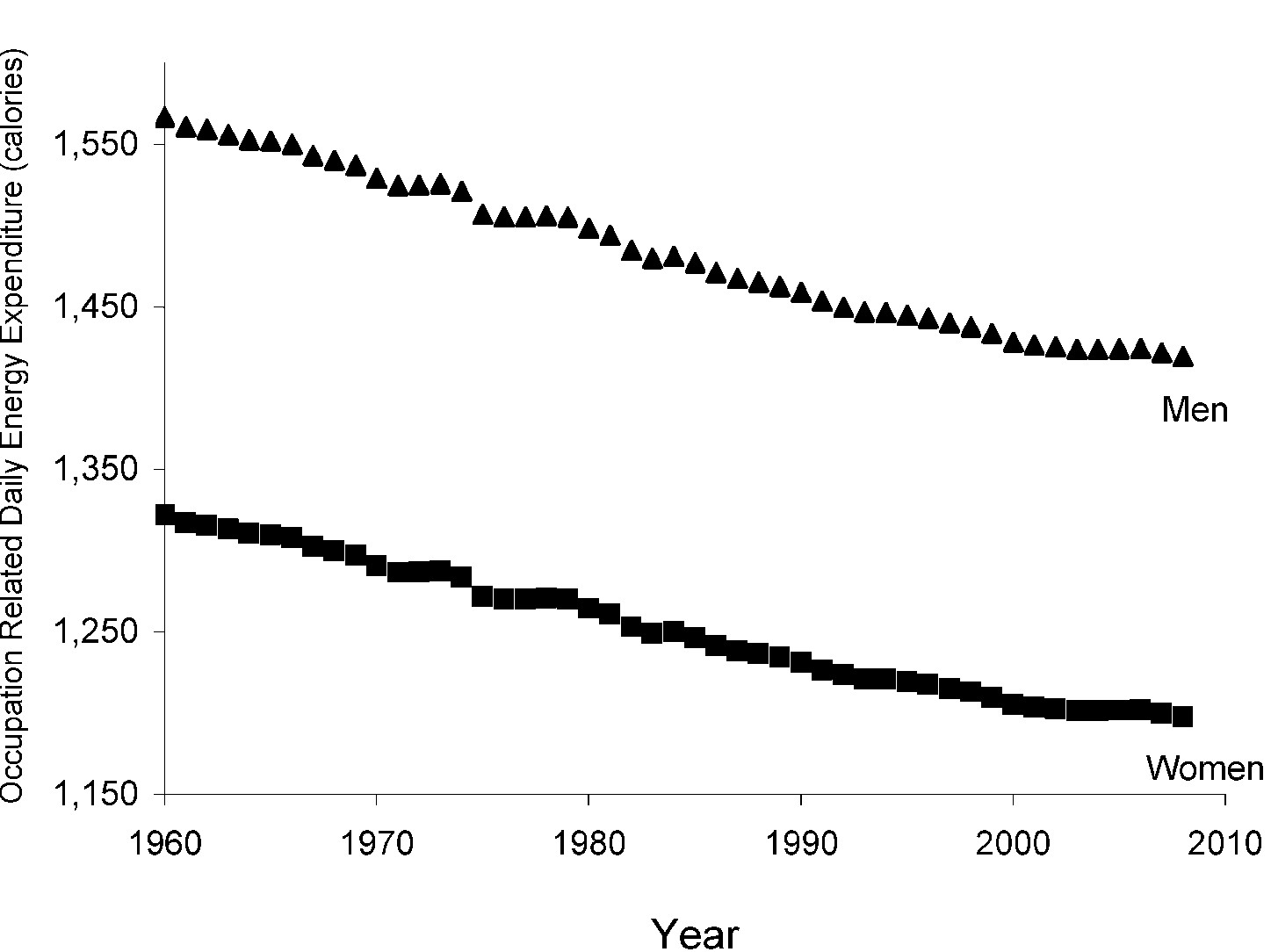
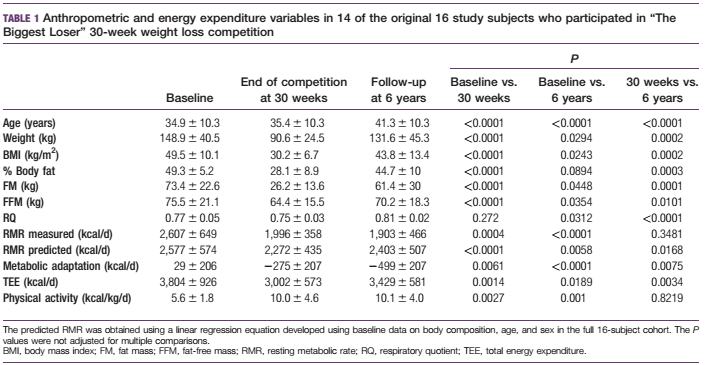
Lihaskasvua käsittelevä artikkelisarja lähenee loppua ja on viimeisen osan vuoro. Tässä artikkelissa käsitellään miten eri yksilöt adaptoituvat voimaharjoitteluun ja miten harjoittelu kannattaisi ohjelmoida, jotta kehitys olisi kaikkein optimaalisinta.
Genetiikan vaikutus lihaskasvuun
Geenit vaikuttavat suuresti henkilön kehittymiseen. Teoreettisesti voidaan ajatella maksimaalisen lihaskasvun rajojen olevan olemassa. Nämä määrittyvät genotyypin, eli geneettisen informaation, ja näiden geenien ilmentymisen yksilössä mukaan. Geenit asettavat teoreettisesti meille tietyt reunaehdot, miten reagoimme erilaisiin tilanteisiin. Geenien takia jotkut yksilöt kehittyvät nopeammin ja jotkut yksilöt hitaammin. Kuiinka paljon vaihtelua voi sitten eri henkilöiden välillä olla?
KUVA 1: Tarkistuslista, jos epäilet salimarkun tai salijonnan käyttävän kehon ulkopuolisia hormoneita.
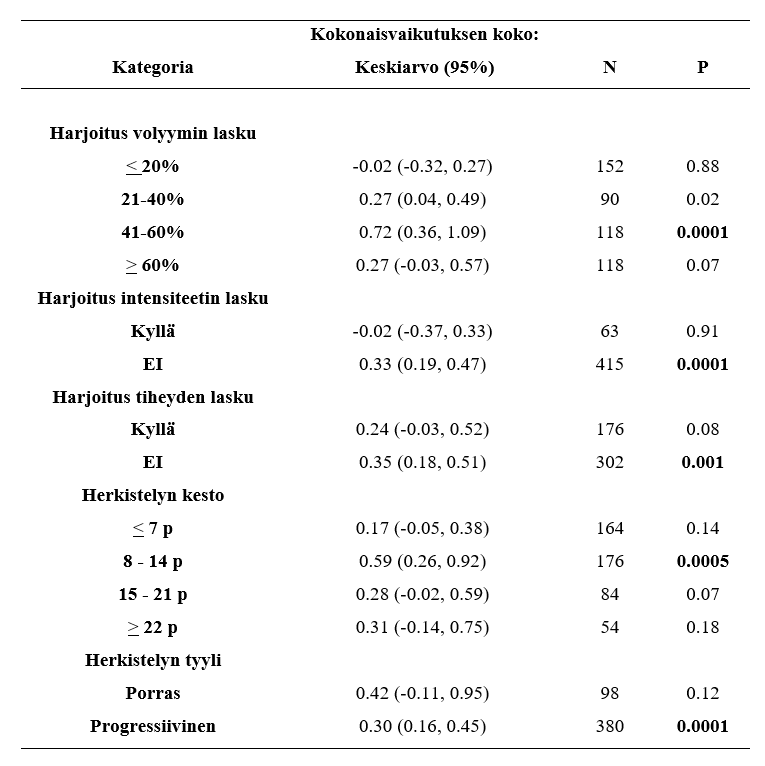
Yksilölliset vasteet harjoitteluun vaihtelevat hurjasti ihmisillä. Tämä tarkoittaa sitä, että jos kaksi samaa ihmistä tekee täysin saman ohjelman niin tuskin koskaan he kehittyvät täsmälleen saman verran. Esimerkiksi samanlaisella harjoittelulla lihaksen kasvu voi vaihdella jopa -11 % - + 30 % välillä ja voima taas - 8 % - + 60 % välillä (Ahtiainen et al., 2016). Ei-respondoivia ihmisiä, eli niitä, jotka eivät reagoi mitenkään tutkimusten voimaharjoitteluun on keskimäärin 25% koehenkilöistä (Bamman et al., 2007). Toisaalta nykyään ajatellaan, että erilainen harjoittelu, ravinto, lepo tai joku muu muuttuja olisi voinut vaikuttaa yksilöllisiin vasteisiin. Eli tälläiset ei-respondoivat koehenkilöt johtuvat pääosin huonosta tutkimusasetelmasta, elämäntilanteesta tai muusta tekijästä, mikä estää kehityksen. Jotkut ei-respondoivista ihmisistä tarvitsevat vain enemmän ja pidempää harjoittelua verrattuna toisiin.
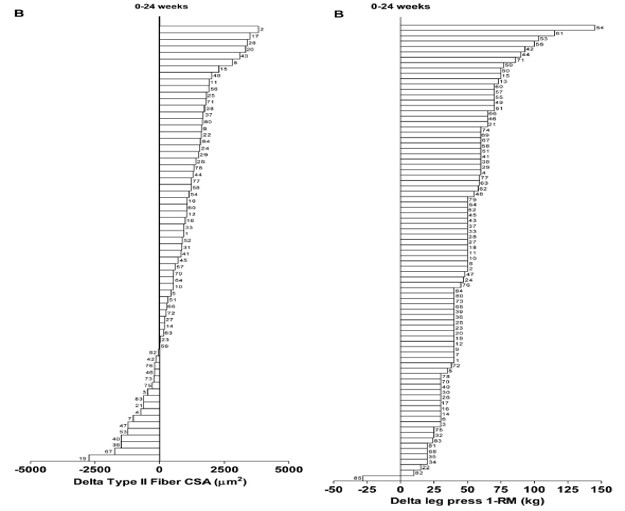
KUVA 2: Oikeassa kaaviossa nähdään, että lähes kaikki koehenkilöt saivat erään tutkimuksen aikana lisää voimaa, mutta vasemmalla huomataan, että joidenkin koehenkilöiden lihaskoko jopa pieneni tutkimuksen aikana. (Churward-Venne et al., 2015).
Mikä näitä eroja sitten selittää? Pescatello ja kummanit (2013) pystyivät tunnistamaan 17 geeniä, joiden uskotaan selittävän osan henkilöiden välisistä eroista. Näistä tärkeimpänä hypertrofian kannalta pidetään tällä hetkellä BMP2-geeniä (Devaney et al., 2009). Ja myös MGF:ää pidetään erittäin tärkeänä (Bamman et al., 2007).
Lisäksi satelliittisolut lisääntyvät ja luovuttavat tumia tehokkaammin hyvin respondoituvilla ihmisillä kuin verrataan ei-respondoituviin ihmisiin (Petrella et al., 2008). Myös lihaksen morfologia vaikuttaa suuresti lihaskasvuun. Mitä enemmän lihassoluja henkilöllä on, niin sen helpommin lihas kasvaa. Lihassolujen määrä on aika vakio ja hyperplasiaa (lihassolujen lisääntymistä) ei tapahdu enää 24 viikon jälkeen syntymästä. Lihassolujen määrä onkin pitkälti geenien määrittämä.
Jos vertaillaan mieskehonrakentajia ja tavallisia kadun tallaajia niin kehonrakentajilla on enemmän lihassoluja hauiksissa ja nämä solut olivat myös isompia verrattuna tavallisiin tallaajiin (MacDougall et al., 1984). Joten niillä, jotka päätyivät kehonrakentajiksi oli jo enemmän lihassoluja kuin muilla normaaleilla ihmisillä. Tämä on yksi syy, miksi kehonrakentajat reagoivat paremmin voimaharjoitteluun.
Nopeiden lihassolujen määrä kasvatettavassa lihaksessa on tärkeä tekijä sen suhteen miten voimaharjoittelu vaikuttaa lihakseen. Tyypin II - lihassolut kasvavat noin 50% enemmän verrattua tyypin I -lihassoluihin. Toisaalta tässäkin suhteessa on hurjasti henkilöiden välistä vaihtelua (Kosek et al., 2006). Tutkimuksia, jotka selittävät fysiologisia mekanismeja yksilöllisessä voimaharjoittelussa on saatavilla erittäin niukasti, mutta erot johtuvat yleisesti perimästä, geenien ilmenemisessä, soluviestinnässä, hormonireseptoreiden toiminnassa ja satelliittisoluissa. Nostan vielä esille ravitsemuksen, jota harvoin tutkimuksissa kontrolloidaan ja sillä on valtava merkitys lihaskasvun mahdollistamisessa.
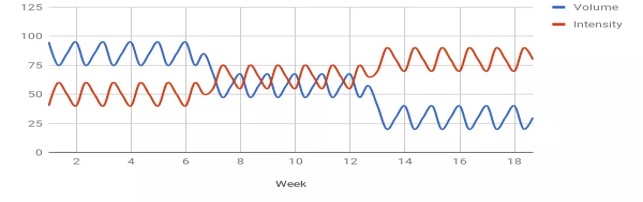
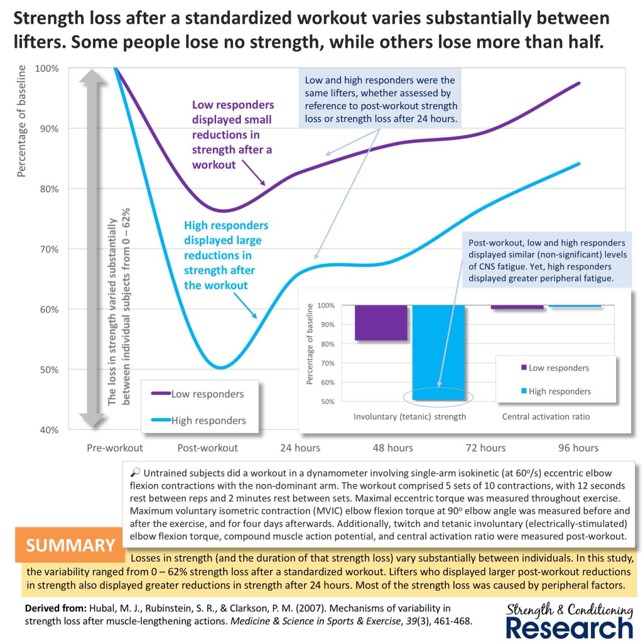
KUVA 3: Näyttäisi myös siltä, että hyvin voimaharjoitteluun reagoivat ihmiset saavat itsestään irti paljon harjoituksen aikana. Heillä harjoituksen jälkeen voima laskee enemmän kuin huonosti respondoituvilla ihmisillä. Tämä tarkoittaa sitä, että he ovat saaneet enemmän väsymystä aikaiseksi harjoituksen aikana.
Genetiikalla on siis kieltämättä iso rooli henkilön lihaskasvussa. Voit kiittää vanhempiasi näistä lahjoista, mutta muuten asiaan ei kannata kiinnittää sen enempää huomiota. Jokainen voi ja pystyy kehittymään, vaikka tutkimuksissa usein löydetään ei-respontoivia henkilöitä harjoitteluun niin tähän saattaa olla syynä vääränlainen tutkimuksenasettelu, harjoittelu tai ohjelmointi. Yksilöllinen harjoittelu ja ohjelma mahdollistavat jokaisen kehittymisen. Heikosti respondoivien pitää joskus reenata jopa paljon vähemmän ja harvemmin kehittyäkseen. Tai kevyemmillä kuormilla. Myös harjoitustausta vaikuttaa suuresti henkilön kykyyn adaptoitua. Aloittelevilla ihmisillä lihaskasvu voi olla jopa 28% ja korkeimmillaan 58% 16 viikossa (Stewart & Rittweger, 2006), kun taas kehonrakentajat kehittyivät viidessä kuukaudessa “vain” 3-7% (Alway et al., 1992). Ohjelmointi ja ohjelman muuttujien manipulointi korostuu erityisesti, kun harjoitustaustaa kertyy enemmän. Kevennetyt jaksot voivat auttaa taas respontoimaan paremmin stimulukseen (Ogasawara et al., 2013).
Ei-respondereilla ei siis ole mielenvika pääsään vaan se on ihan oikea asia. Heidän tulee erityisesti keskittyä ravintoon ja oikeanlaiseen harjoitteluun. Huonosti respondoivat saattavat olla vain hitaammin respondoivia ja tarvitsevat enemmän harjoitusvolyymia kehittyäkseen. Nimi on vähän harhaanjohtava sillä jokainen kehittyy oikeanlaisella harjoittelulla. Saatat vain olla heikommin ja hitaammin respondoituva henkilö verrattuna salikaveriisi. Lisäksi vielä useissa tutkimuksissa kehitys on 12 viikossa esim. non respondereilla “vain” 5%. Tämä tulos luokitellaan tutkimuksissa hitaasti reaspondoivaksi, mutta jos kehitys jatkuisi koko vuoden verran samanlaisena se vastaisi alle vuodessa esimerkiksi 100kg penkissä siirtymistä 121,6 kiloon. Kuinka moni olisi tyytyväinen tuohon kehitykseen vuodessa? Minä ainakin olisin. Joten nimi on hiukan harhaanjohtava ja yleisesti ongelmat johtuvat tutkimusasetelmasta, sekä heterogeenisestä tutkimusjoukosta.
Neuvostoliitossa
periodisaation (ja ylipäätään liikuntatieteen) tutkimus- ja kehitystyö oli
tuolloin huomattavan laajaa ja systemaattista. Tämän tutkimustyön tuloksena
syntyi klassinen periodisaatiomalli. Muissakin maissa toki käytettiin ja
kehitettiin samantyylistä periodisaatiota, tosin pienemmässä mittakaavassa.
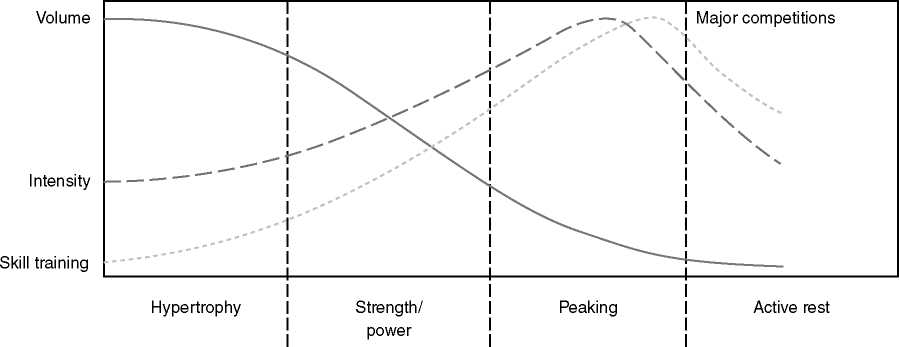
KUVA 4: Klassinen periodisaatiomalli, missä harjoitusvolyymi on aluksi korkealla ja vähenee pääkilpailun lähestyessä. Samalla harjoittelun intensiteetti nousee.
Periodisaatio
on erittäin vähän tutkittu aihealue. Johtuen luultavasti siitä, että
periodisaatio ei ole mikään selkeästi käsitettävä kokonaisuus. Erillaisia
malleja voidaan esittää, mutta näidenkin sisällä voi olla loputtamattomasti
erillaisia muutujia, joita voi siirrellä ja muutella. Periodisaatio perustuu
Selyen yleiseeen adaptaatio syndrooma - teoriaan (GAS - general adaptation
syndrome). Se perustuu kehon reaktioihin, jotka ovat hälytys, vastustus ja
uupuminen. (Selye, 1950). Teoriaa on nykypäivinä kritisoitu
yksinkertaistamisesta ja siitä, että se ei huomioi esimerkiksi psykologisia,
teknisiä ja ravinnollisia seikkoja. Selyen teorian mukaan harjoittelua
pidetään stressitekijänä, johon voidaan soveltaa GAS - teoriaa. Harjoittelun
tulee olla progressiivisesti haastavaa, jotta tasannevaihetta ei tule
harjoittelussa ja toisaalta harjoittelu ei saa olla ylikuluttavaa.
Periodisaatiomalleista kolmea on tutkittu lihashypertrofian kannalta: perinteinen lineaarinen malli, ei-lineaarinen ja käänteinen periodisaatio.
Perinteisessä
lineaarisessa mallissa on vähintään makrosykli, mesosykli ja mikrosykli
(esimerkiksi vuosi, peruskuntokausi 8 vko, yksi harjoitusviikko). Malli etenee
korkeasta voluumista ja matalasta intensiteetistä kohti korkeaan
intensiteettiin ja matalaan voluumiin. Esimerkiksi rakenteellinen
jakso/hypertrofinen jakso, jossa voluumi nousee ja intensiteetti pysyy 60-75%
välillä, seuraavaksi voi olla perusvoimakausi intensiteetin ollessa 80-90%
välillä ja voluumin vähetessä. Viimeinen mesosykli olisi intensiteetiltään
suurin 90-100%, mutta voluumiltaan pienin.
Lihaskasvun osalta aihetta on tutkittu jonkin verran, mutta itse pidän tutkimuksia metodologisilta osuuksilta ontuvilta, joten johtopäätöksiä ei tule vetää vielä näiden tutkimusten pohjalta. Stone ja kumppanit (1981) tutkivat periodisaatioidun ohjelman ja ei-periodisoidun ohjelman eroja, ja huomasivat hypertrofian (mitattu veden alaispunnituksella) eroavan tilastollisesti merkitsevästi ei-periodisoidusta ohjelmoinnista. Baker:in tutkimusporukka taa (1994) ei löytänyt eroja periodisatioidun ja ei-periodisaatiouidun välille (ihopoimumittaus). Kun taas Monteiro ja kumppanit (2009) löysivät eron (ihopoimu, ei tilastollisesti merkittävä) periodisoidun harjoittelun hyväksi.
Voiman osalta periodisoitu harjoittelu tuottaa kiistatta parempia tuloksia kuin ei periodisoitu harjoittelu (Ahmadizad et al., 2014;, Monteiro et al., 2009,Willoughby, 1993). Voisi ajatellakin, että koska voima nousee paremmin periodisoidulla harjoittelulla, johtaisi se mekaanisen kuormituksen johdosta myös suurempiin hypertrofisiin adaptaatioihin ajan kanssa, mutta tätä ei ole vielä todistettu.
KUVA 5: Periodisoitu harjoittelu tuottaa keskimäärin 21,78 – 23,62% voiman nousun, kun taas ei periodisoitu harjoittelu johtaa vain 18,90 – 19,10% keskimääräiseen voiman nousuun. Viikkotasolla samat luvut 1,96 % - 2,05 % verrattuna ei periodisoituun 1,59 – 1,70%. (Nuckols, 2018.)
Ei-lineaarinen periodisaatio tai aaltoileva periodisaatio on nykyään yksi suosituimmista suuntauksista. Sitä on käytetty jo kauan, mutta Poliquin (1988) oli ensimmäinen, joka esitteli sen tiedeyhteisössä. Hänen mukaansa lineaarisen ohjelmoinnin ongelmat ovat liian suuri progressiivinen painon lisäys liian lyhyessä ajassa ja alussa saavutettujen hypertrofisten adaptaatioiden menetys muissa mesosykleissä, koska voluumi laskee liikaa.
Aaltoilevassa periodisaatiossa vaihdellaan voluumia ja intensiteettiä kuinkas muutenkaan kuin aaltoilevaan tyyliin. Mallia on kehitetty eteenpäin ja tällä hetkellä suosituin malli taitaa olla päivittäin vaihtuva periodisaatio (daily undulating periodization - DUP). Tässä mallissa on viikon sisällä maksimivoima, perusvoima ja hypertrofinen harjoitus. Aihetta on tutkittu suhteellisen laajasti. Ja vain yksi tutkimus sai tilastollisesti merkitsevän eron lineaarisen ja ei-lineaarisen mallin välille. Tässä tutkimuksessa ei-lineaarinen malli oli selvästi parempi lisäämään kyynärpään koukistajien ja ojentajien tiheyttä (Simao et al., 2012). Muuten muissa tutkimuksissa ei ole saatu eroja mallien välille lihaskasvun osalta, mikä osoittaa kummankin mallin olevan yhtä toimiva lihaskasvun osalta (Baker et al., 1994; Davitt et al., 2014; , Harries et al., 2015; Monteiro et al., 2009, Prestes et al., 2009, Simao et al., 2012). Toisaalta tutkimusten kesto on ollut korkeimmillaan vain 12 viikkoa ja kun puhutaan periodisaatiosta yleisesti urheilijoiden osalta puhutaan vuosista.
Itse suosin valmennettavillani enemmän hybridimallia, joka on versio aaltoilevasta mallista. Tässä mallissa harjoitusohjelma on laadittu blokkeihin ja näiden blokkien sisällä tapahtuu aaltoilua. Blokit toisaalta etenevät lineaarisesti. Viikon sisällä siinä tehdään maksimivoimasarjoja ja perusvoima sarjoja samassa harjoituksessa järkevästi ohjelmoiden. Pitkillä perusvoimakausilla saattaa hävitä liikaa maksimivoimaa ja pitkillä maksimivoima kausilla saattaa hävitä liikaa lihasta tai perusvoimaa, joten järkevä hybridiohjelmointi poistaa nämä ongelmat.
KUVA 6: Aaltoileva periodisaatio, missä volyymi ja intensiteetti aaltoilevat mahdollistaen intensiteetin ylläpidon peruskuntokaudella ja volyymin ylläpidon kisakaudella.
Käänteistä periodisaatiota on ehdotettu lihaskasvuun tähtääville tärkeimmäksi ohjelmaksi. Lineaarinen malli tiputtaa jatkuvasti voluumia kohti kauden päätähtäintä ja kun tiedetään voluumin olevan elintärkeä hypertrofian kannalta voisi ajatella lineaarisen mallin olevan huono lihaskasvun kannalta. Käänteisessä mallissa voluuumi on korkeimmillaan kauden päätähtäimen kohdalla. Aihe on vielä tutkimuskentällä tuore ja vain tietääkseni Prestes ja kumppanit (2009) ovat sitä tutkineet huonoin tuloksin käänteisen mallin puolesta. Lineaarinen malli oli huomattavasti parempi kuin käänteinen malli. Toisaalta mittauksissa käytettiin ihopoimupihtejä, jotka ovat metodisesti yhtä tyhjän kanssa.
Tiettyä suunnitelmallisuutta harjoitteluun tarvitaan, koska muuten harjoittelusta puuttuu variaatio, progressiivisuus ja tavoitteet. Lihaskasvun osalta yksi tärkeimmistä periodisaation tuomista asioista on kevennetyt jaksot. Keho ottaa paremmin vastaan harjoittelua kuin välillä tiputtaa voluumia ja antaa kropan palautua. Lisäksi on tärkeä kerätä palautetta periodisaation toimivuudesta ja suunnitelman muokkaaminen sen mukaan auttavat kehittämään ohjelmointia.
Kiitos lukijalle, jos jaksoit lukea tänne saakka! Alla lähdeluettelo kaikista artikkelisarjan osista. Aiheeseen liittyvä lukeminen ei ihan heti lopu kesken. Jätä palautetta tai kommentteja ja tsemppiä treeneihin!
Lähteet:
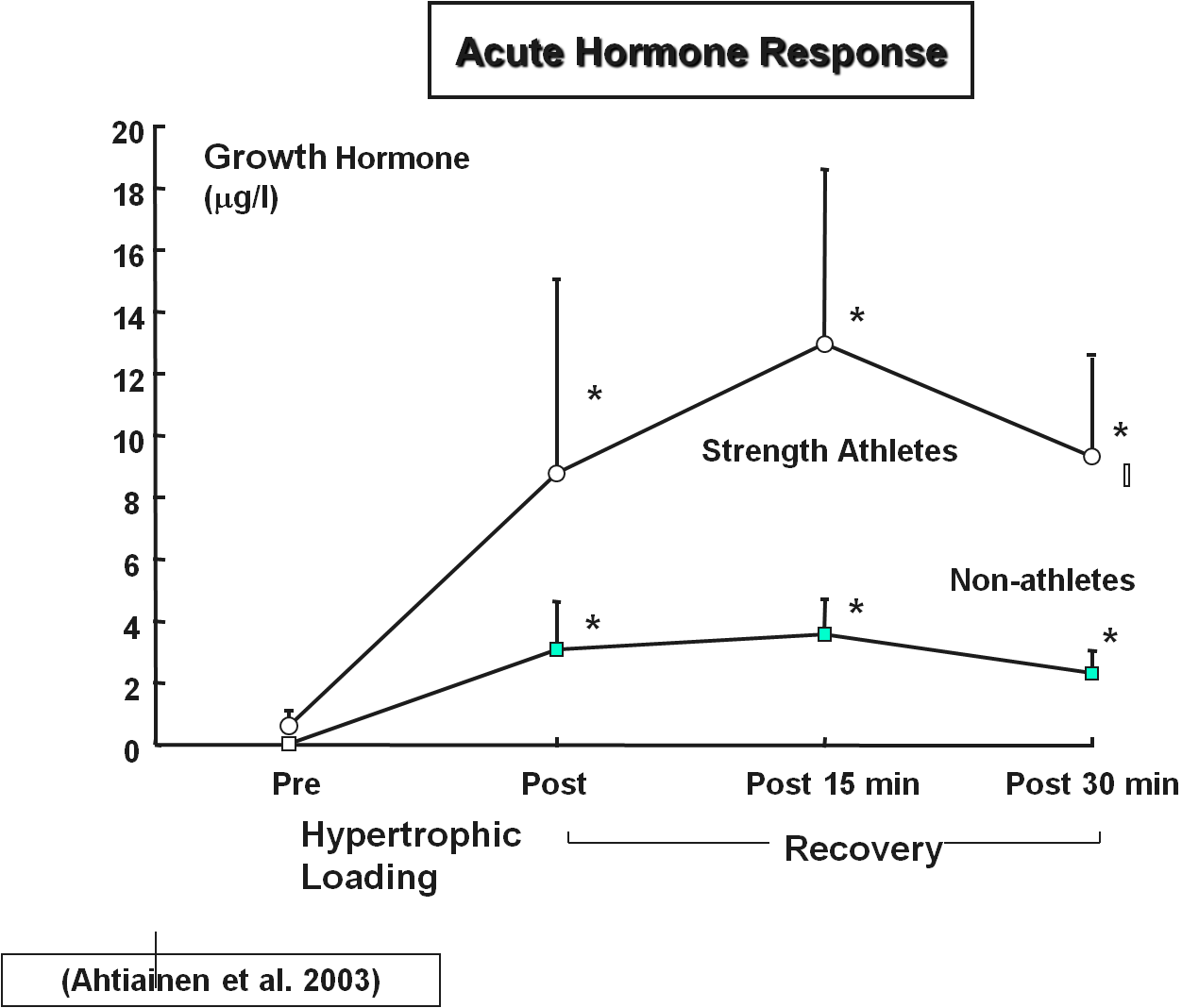
Ahtiainen, J., Pakarinen, A., Kraemer, W. &
Häkkinen, K. (2003). Acute
Hormonal and Neuromuscular Responses and Recovery to Forced vs. Maximum
Repetitions Multiple Resistance Exercises. International Journal of Sports
Medicine, 24(6), 410–418. doi: 10.1055/s-2003-41171
Ahtiainen, J. P.,
Pakarinen, A., Alen, M., Kraemer, W. J., & Häkkinen, K. (2005). Short vs. Long Rest Period Between the Sets in
Hypertrophic Resistance Training: Influence on Muscle Strength, Size, and
Hormonal Adaptations in Trained Men. The Journal of Strength and Conditioning
Research, 19(3), 572. doi: 10.1519/15604.1
Ahtiainen JP, Walker S, Peltonen H, Holviala J,
Sillanpää E, Karavirta L, et al. Heterogeneity in resistance
training-induced muscle strength and mass responses in men and women of
different ages. Age (Omaha). 2016;38:1–13.
Ahmadizad, S., Ghorbani, S., Ghasemikaram, M. &
Bahmanzadeh, M. (2014) Effects of short-term nonperiodized, linear periodized
and daily undulating periodized resistance training on plasma adiponectin,
leptin and insulin resistance. Clinical biochemistry, 47: 417-422.
Alegre, L.,
Jimenez, F., Gonzalo-Orden, J., Acero, R. & Aguado, X. (2006). Effects of
dynamic resistance training on fascicle length and isometric strength. Journal
of sports sciences: 24. 501-8. 10.1080/02640410500189322.
Allen, D. G.,
Whitehead, N. P., & Yeung, E. W. (2005). Mechanisms of stretch-induced
muscle damage in normal and dystrophic muscle: role of ionic changes. The
Journal of Physiology, 567(3), 723–735. doi: 10.1113/jphysiol.2005.091694
Alway, S. E.,
Grumbt, W. H., Stray-Gundersen, J., & Gonyea, W. J. (1992). Effects of
resistance training on elbow flexors of highly competitive bodybuilders.
Journal of Applied Physiology, 72(4), 1512–1521. doi:
10.1152/jappl.1992.72.4.1512
Atherton, P. J.,
Babraj, J., Smith, K., Singh, J., Rennie, M. J., & Wackerhage, H. (2005).
Selective activation of AMPK-PGC-1α or PKB-TSC2-mTOR signaling can explain
specific adaptive responses to endurance or resistance training-like electrical
muscle stimulation. The FASEB Journal, 19(7), 786–788. doi:
10.1096/fj.04-2179fje
Baker, D., Wilson, G. &
Carolyn, R. (1994) Periodization: The effect on strength of manipulating volume
and intensity. Journal of strength and conditioning research, 8: 235-242.
Bamman, M. M.,
Petrella, J. K., Kim, J.-S., Mayhew, D. L., & Cross, J. M. (2007). Cluster analysis tests the importance of myogenic gene
expression during myofiber hypertrophy in humans. Journal of Applied
Physiology, 102(6), 2232–2239. doi: 10.1152/japplphysiol.00024.2007
Benziane, B.,
Burton, T. J., Scanlan, B., Galuska, D., Canny, B. J., Chibalin, A. V., …
Stepto, N. K. (2008). Divergent cell signaling after short-term intensified
endurance training in human skeletal muscle. American Journal of
Physiology-Endocrinology and Metabolism, 295(6). doi:
10.1152/ajpendo.90428.2008
Blazevich, A. J.,
Cannavan, D., Coleman, D. R., & Horne, S. (2007). Influence of concentric
and eccentric resistance training on architectural adaptation in human
quadriceps muscles. Journal of Applied Physiology, 103(5), 1565–1575. doi:
10.1152/japplphysiol.00578.2007
Bondesen, B. A.,
Mills, S. T., & Pavlath, G. K. (2006). The COX-2 pathway regulates growth
of atrophied muscle via multiple mechanisms. American Journal of
Physiology-Cell Physiology, 290(6). doi: 10.1152/ajpcell.00518.2005
Bondesen, B. A.,
Mills, S. T., Kegley, K. M., & Pavlath, G. K. (2004). The COX-2 pathway is
essential during early stages of skeletal muscle regeneration. American Journal
of Physiology-Cell Physiology, 287(2). doi: 10.1152/ajpcell.00088.2004
Brentano, M. &
Martins Kruel, L. (2011) A review on strength exercise-induced muscle damage:
applications, adaption mechanisms and limitations. J Sports Med Phys Fitness,
51(1): 1-10.
Burd, N. A.,
Holwerda, A. M., Selby, K. C., West, D. W. D., Staples, A. W., Cain, N. E., …
Phillips, S. M. (2010). Resistance exercise volume affects myofibrillar protein
synthesis and anabolic signalling molecule phosphorylation in young men. The
Journal of Physiology, 588(16), 3119–3130. doi: 10.1113/jphysiol.2010.192856
Burd, N. A.,
Mitchell, C. J., Churchward-Venne, T. A., & Phillips, S. M. (2012). Bigger
weights may not beget bigger muscles: evidence from acute muscle protein
synthetic responses after resistance exercise. Applied Physiology, Nutrition,
and Metabolism, 37(3), 551–554. doi: 10.1139/h2012-022
Choi J., Takahashi H., Itai Y. (1998) The
difference between effects of ‘power-up type’ and ‘bulk-up type’ strength
training exercises: With special reference to muscle cross-sectional area. Jpn J Phys Fitness Sports Med 47(1), 119-129.
Clarkson, P. M.,
Byrnes, W. C., Mccormick, K. M., Turcotte, L. P., & White, J. S. (1985).
Muscle Soreness And Serum Creatine Kinase Activity Following Isometric,
Eccentric And Concentric Exercise. Medicine & Science in Sports &
Exercise, 17(2), 277. doi: 10.1249/00005768-198504000-00424
Coffey, V. G.,
Zhong, Z., Shield, A., Canny, B. J., Chibalin, A. V., Zierath, J. R., &
Hawley, J. A. (2006). Early signaling responses to divergent exercise stimuli
in skeletal muscle from well-trained humans. The FASEB Journal, 20(1), 190–192.
doi: 10.1096/fj.05-4809fje
Cornelison, D.
& Wold, B. (1997) Single-cell analysis of regulatory gene expression in
quiescent and activated mouse skeletal muscle satellite cells. Advances in
developmental biology, 191(2): 270–283.
Conboy, I. M., Conboy, M. J.,
Wagers, A. J., Girma, E. R., Weissman, I. L., & Rando, T. A. (2005).
Rejuvenation of aged progenitor cells by exposure to a young systemic
environment. Nature, 433(7027): 760–764. doi: 10.1038/nature03260
Dangott, B., Schultz, E.,
& Mozdziak, P. E. (2000). Dietary Creatine Monohydrate Supplementation
Increases Satellite Cell Mitotic Activity During Compensatory Hypertrophy.
International Journal of Sports Medicine, 21(1), 13–16. doi:
10.1055/s-2000-8848
Davitt, P. M., Pellegrino, J.
K., Schanzer, J. R., Tjionas, H., & Arent, S. M. (2014). The Effects of a
Combined Resistance Training and Endurance Exercise Program in Inactive College
Female Subjects. Journal of Strength and Conditioning Research, 28(7),
1937–1945. doi: 10.1519/jsc.0000000000000355
Devaney, J. M., Tosi, L. L.,
Fritz, D. T., Gordish-Dressman, H. A., Jiang, S., Orkunoglu-Suer, F. E., …
Rogers, M. B. (2009). Differences in fat and muscle mass associated with a
functional human polymorphism in a post-transcriptionalBMP2gene regulatory
element. Journal of Cellular Biochemistry, 107(6), 1073–1082. doi:
10.1002/jcb.22209
Dhawan, J., & Rando, T. A.
(2005). Stem cells in postnatal myogenesis: molecular mechanisms of satellite
cell quiescence, activation and replenishment. Trends in Cell Biology, 15(12),
666–673. doi: 10.1016/j.tcb.2005.10.007
Egner, I. M., Bruusgaard, J.
C., Eftestøl, E., & Gundersen, K. (2013). A cellular memory mechanism aids
overload hypertrophy in muscle long after an episodic exposure to anabolic
steroids. The Journal of Physiology, 591(24): 6221–6230. doi:
10.1113/jphysiol.2013.264457
Evans, W. J., & Cannon, J.
G. (1991). The Metabolic Effects of Exercise-Induced Muscle Damage. Exercise
and Sport Sciences Reviews, 19(1). doi: 10.1249/00003677-199101000-00003
Finkenzeller, G., Newsome, W.,
Lang, F., & Häussinger, D. (1994). Increase of c-jun mRNA upon hypo-osmotic
cell swelling of rat hepatoma cells. FEBS Letters, 340(3), 163–166. doi:
10.1016/0014-5793(94)80129-0
Folland, J. &
Williams, A. (2007) The adaptations to strength training - morphological and
neurological contributions to increased strength. Sports medicine, 37 (2):
145-168.
Fonseca, R. M.,
Roschel, H., Tricoli, V., Souza, E. O. D., Wilson, J. M., Laurentino, G. C., …
Ugrinowitsch, C. (2014). Changes in Exercises Are More Effective Than in
Loading Schemes to Improve Muscle Strength. Journal of Strength and
Conditioning Research, 28(11), 3085–3092. doi: 10.1519/jsc.0000000000000539
Fry, C. S., Glynn,
E. L., Drummond, M. J., Timmerman, K. L., Fujita, S., Abe, T., … Rasmussen, B.
B. (2010). Blood flow restriction exercise stimulates mTORC1 signaling and
muscle protein synthesis in older men. Journal of Applied Physiology, 108(5),
1199–1209. doi: 10.1152/japplphysiol.01266.2009
Gardiner NE. Athletics of the ancient world. Oxford:
University Press; 1930.
Glass, D. J.
(2010). PI3 Kinase Regulation of Skeletal Muscle Hypertrophy and Atrophy.
Current Topics in Microbiology and Immunology Phosphoinositide 3-Kinase in
Health and Disease, 267–278. doi: 10.1007/82_2010_78
Goodman, C. A.,
Mayhew, D. L., & Hornberger, T. A. (2011). Recent progress toward
understanding the molecular mechanisms that regulate skeletal muscle mass.
Cellular Signalling, 23(12), 1896–1906. doi: 10.1016/j.cellsig.2011.07.013
Gordon, S. E.,
Kraemer, W. J., Vos, N. H., Lynch, J. M., & Knuttgen, H. G. (1994). Effect
of acid-base balance on the growth hormone response to acute high-intensity
cycle exercise. Journal of Applied Physiology, 76(2), 821–829. doi:
10.1152/jappl.1994.76.2.821
Goto, K., Ishii,
N., Kizuka, T. & Takamatsu, K. (2005) The impact of metabolic stress on
hormonal responses and muscular adaptations. Medicine and science in sports and
exercise, 37(6): 955-963.
Guyton. A. C., Hall J. E. 2011. Textbook of Medical
Physiology. 12th edition. Philadelphia, USA : W.B. Saunders Company
Harries, S. K.,
Lubans, D. R., & Callister, R. (2016). Comparison of resistance training
progression models on maximal strength in sub-elite adolescent rugby union
players. Journal of Science and Medicine in Sport, 19(2), 163–169. doi:
10.1016/j.jsams.2015.01.007
Hackett, D. A.,
Johnson, N. A., & Chow, C.-M. (2013). Training Practices and Ergogenic Aids
Used by Male Bodybuilders. Journal of Strength and Conditioning Research,
27(6), 1609–1617. doi: 10.1519/jsc.0b013e318271272a
Handayaningsih,
A.-E., Iguchi, G., Fukuoka, H., Nishizawa, H., Takahashi, M., Yamamoto, M., …
Takahashi, Y. (2011). Reactive Oxygen Species Play an Essential Role in IGF-I
Signaling and IGF-I-Induced Myocyte Hypertrophy in C2C12 Myocytes.
Endocrinology, 152(3), 912–921. doi: 10.1210/en.2010-0981
Harber, M. P.,
Konopka, A. R., Douglass, M. D., Minchev, K., Kaminsky, L. A., Trappe, T. A.,
& Trappe, S. (2009). Aerobic exercise training improves whole muscle and
single myofiber size and function in older women. American Journal of
Physiology-Regulatory, Integrative and Comparative Physiology, 297(5). doi:
10.1152/ajpregu.00354.2009
Harber, M. P.,
Konopka, A. R., Undem, M. K., Hinkley, J. M., Minchev, K., Kaminsky, L. A., …
Trappe, S. (2012). Aerobic exercise training induces skeletal muscle
hypertrophy and age-dependent adaptations in myofiber function in young and
older men. Journal of Applied Physiology, 113(9), 1495–1504. doi:
10.1152/japplphysiol.00786.2012
Harridge, S.
(2007) Plasticity of human skeletal muscle: Gene expression to in vivo
function. Exp Physiolo. 92: 738-797.
Helms, E., Aragon,
A. & Fitschen, P. (2014). Evidence-based recommendations for natural
bodybuilding contest preparation: Nutrition and supplementation. Journal of the
International Society of Sports Nutrition. 11. 20. 10.1186/1550-2783-11-20.
Hill, M., Wernig,
A., & Goldspink, G. (2003). Muscle satellite (stem) cell activation during
local tissue injury and repair. Journal of Anatomy, 203(1), 89–99. doi:
10.1046/j.1469-7580.2003.00195.x
Hornberger, T. A.,
Chu, W. K., Mak, Y. W., Hsiung, J. W., Huang, S. A., & Chien, S. (2006).
The role of phospholipase D and phosphatidic acid in the mechanical activation
of mTOR signaling in skeletal muscle. Proceedings of the National Academy of Sciences,
103(12), 4741–4746. doi: 10.1073/pnas.0600678103
Izquierdo, M.,
Ibañez, J., González-Badillo, J. J., Häkkinen, K., Ratamess, N. A., Kraemer, W.
J., French, D., Eslava, J., Altadill, A., Asiain X. & Gorostiaga, E. M.
(2006). Differential effects of strength training leading to failure versus not
to failure on hormonal responses, strength, and muscle power gains. Journal of
Applied Physiology, 100(5), 1647–1656. doi: 10.1152/japplphysiol.01400.2005
Jacinto, E., &
Hall, M. N. (2003). TOR signalling in bugs, brain and brawn. Nature Reviews
Molecular Cell Biology, 4(2), 117–126. doi: 10.1038/nrm1018
Kadi, F.,
Eriksson, A., Holmner, S., Butler-Browne, G. & Thornell, L. (1999) Cellular
adaptation of the trapezius muscle in strength-trained athletes. Histochemistry
and cell biology, 111(3): 189-95.
Kadi, F. &
Thornell, L.E. (2000) Concomitant increases in myonuclear and satellite cell
content of female trapezius muscle following strength training. Histochemistry
and cell biology 113, 99-103.
Keogh, J. W.,
Wilson, G. J., & Weatherby, R. E. (1999). A Cross-Sectional Comparison of
Different Resistance Training Techniques in the Bench Press. Journal of
Strength and Conditioning Research, 13(3), 247–258. doi:
10.1519/00124278-199908000-00012
Kohn, T. A., Essén-Gustavsson,
B., & Myburgh, K. H. (2010). Specific muscle adaptations in type II fibers
after high-intensity interval training of well-trained runners. Scandinavian
Journal of Medicine & Science in Sports, 21(6), 765–772. doi:
10.1111/j.1600-0838.2010.01136.x
Kosek, D. J., Kim,
J.-S., Petrella, J. K., Cross, J. M., & Bamman, M. M. (2006). Efficacy of 3
days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in
young vs. older adults. Journal of Applied Physiology, 101(2), 531–544. doi:
10.1152/japplphysiol.01474.2005
Krieger, J. W.
(2010). Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy:
A Meta-Analysis. Journal of Strength and Conditioning Research, 24(4),
1150–1159. doi: 10.1519/jsc.0b013e3181d4d436
Lang, F (2007).
Mechanisms and Significance of Cell Volume Regulation. J Am Coll Nutr., 26:
613s-623s.
Lang, F., Busch,
G. L., Ritter, M., Völkl, H., Waldegger, S., Gulbins, E., & Häussinger, D.
(1998). Functional Significance of Cell Volume Regulatory Mechanisms.
Physiological Reviews, 78(1), 247–306. doi: 10.1152/physrev.1998.78.1.247
Loenneke, J. P.,
Wilson, J. M., Marín, P. J., Zourdos, M. C., & Bemben, M. G. (2011). Low
intensity blood flow restriction training: a meta-analysis. European Journal of
Applied Physiology, 112(5), 1849–1859. doi: 10.1007/s00421-011-2167-x
Loenneke, J. P.,
Thiebaud, R. S., & Abe, T. (2014). Does blood flow restriction result in
skeletal muscle damage? A critical review of available evidence. Scandinavian
Journal of Medicine & Science in Sports, 24(6). doi: 10.1111/sms.12210
Lynn, R. &
Morgan, D. (1994) Decline running produces more sarcomeres in rat vastus
intermedius muscle fibers than does incline running. Journal of applied
physiology: 77: 1439-1444.
Malm, C. (2001).
Exercise-induced muscle damage and inflammation: fact or fiction? Acta
Physiologica Scandinavica, 171(3), 233–239. doi:
10.1046/j.1365-201x.2001.00825.x
Macdougall, J. D.,
Gibala, M., Tarnopolsky, M., Macdonald, J., Interisano, S., & Yarasheski,
K. (1995). The Time Course For Elevated Muscle Protein Synthesis Following
Heavy Resistance Exercise. Medicine & Science in Sports & Exercise,
27(Supplement). doi: 10.1249/00005768-199505001-00367
MacDougall, D. Ward, G., Sale, D. & Sutton, J.
(1977) Biochemical adaptation of human skeletal muscle to heavy resistance
training and immobilization. Journal of applied physiology:
respiratory, environmental and exercise physiology: 43(4): 700-3.
Macdougall, J. D., Sale, D. G., Alway, S. E., &
Sutton, J. R. (1984). Muscle fiber number in biceps brachii in bodybuilders and
control subjects. Journal of Applied Physiology, 57(5), 1399–1403. doi:
10.1152/jappl.1984.57.5.1399
Mascher, H., Andersson, H., Nilsson, P.-A., Ekblom,
B., & Blomstrand, E. (2007). Changes in signalling pathways regulating
protein synthesis in human muscle in the recovery period after endurance
exercise. Acta Physiologica, 191(1), 67–75. doi: 10.1111/j.1748-1716.2007.01712.x
Masuda K., Choi J.Y., Shimojo H., Katsuta S. (1999) Maintenance of myoglobin concentration in human skeletal muscle
after heavy resistance training. European
Journal of Applied Physiology and Occupational Physiology 79(4), 347-352. [PubMed] [Google Scholar]
Martineau, L. & Gardiner,
P. (2002) Skeletal muscle is sensitive to the tension-time integral but not to
the rate of change of tension, as assessed by mechanically induced signaling.
Journal of biomechanic, 35: 657-663.
Mchugh, M. P. (2003). Recent advances in the
understanding of the repeated bout effect: the protective effect against muscle
damage from a single bout of eccentric exercise. Scandinavian Journal of
Medicine and Science in Sports, 13(2), 88–97. doi:
10.1034/j.1600-0838.2003.02477.x
Mcpherron, A. C., Lawler, A.
M., & Lee, S.-J. (1997). Regulation of skeletal muscle mass in mice by a
new TGF-p superfamily member. Nature, 387(6628), 83–90. doi:
10.1038/387083a0
Mikkola,
J., Rusko, H., Izquierdo, M., Gorostiaga, E., & Häkkinen, K. (2012). Neuromuscular and Cardiovascular Adaptations During
Concurrent Strength and Endurance Training in Untrained Men. International
Journal of Sports Medicine, 33(09), 702–710. doi: 10.1055/s-0031-1295475
Moore, D., Phillips, S., Babraj, J., Smith, K. & Rennie, M. (2005) Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. American journal of physiology - endocrinology andmetabolism, 288: E1153–E1159.
Monteiro, A., Aoki., M.,
Evangelista,A., Alveno, D., Monteiro, G., Picarro Ida, C. & Ugrinowitcch,
C. (2009) Nonlinear periodization maximizes strength gains in split resistance
training routines. Journal of strength and conditioning research, 23: 1321-1326.
Netreba, A., Popov, D., Bravyy,
Y., Lyubaeva, E., Terada, M., Ohira, T.,
Okabe, H., Vinogradova, O.& Ohira, Y. (2013). Responses of knee extensor
muscles to leg press training of various types in human. Rossiĭskii
fiziologicheskiĭ zhurnal imeni I.M. Sechenova / Rossiĭskaia akademiia nauk. 99.
406-16.
Nguyen, H. X., & Tidball,
J. G. (2003). Null Mutation of gp91phoxReduces Muscle Membrane Lysis During
Muscle Inflammation in Mice. The Journal of Physiology, 553(3), 833–841. doi:
10.1113/jphysiol.2003.051912
Nosaka, K., Lavender, A.,
Newton, M. & Sacco, P. (2003) Muscle damage in resistance training – is muscle
damage necessary for strength gain and muscle hypertrophy? International
journal of sport and health science, 1(1):1-8.
Nuckols, G. (2018)
Periodization: What the data say. https://www.strongerbyscience.com/periodization-data/
Ogasawara, R., Kobayashi, K.,
Tsutaki, A., Lee, K., Abe, T., Fujita, S., … Ishii, N. (2013). mTOR signaling
response to resistance exercise is altered by chronic resistance training and
detraining in skeletal muscle. Journal of Applied Physiology, 114(7), 934–940.
doi: 10.1152/japplphysiol.01161.2012
Pescatello, L. S., Devaney, J.
M., Hubal, M. J., Thompson, P. D., & Hoffman, E. P. (2013). Highlights from
the Functional Single Nucleotide Polymorphisms Associated with Human Muscle
Size and Strength or FAMuSS Study. BioMed Research International, 2013, 1–11.
doi: 10.1155/2013/643575
Petrella, J. K., Kim, J.-S.,
Mayhew, D. L., Cross, J. M., & Bamman, M. M. (2008). Potent myofiber
hypertrophy during resistance training in humans is associated with satellite
cell-mediated myonuclear addition: a cluster analysis. Journal of Applied
Physiology, 104(6), 1736–1742. doi: 10.1152/japplphysiol.01215.2007
Pinto, R., Gomes, N.,
Radaelli, R., Botton, C., Brown, L. & Bottaro, M. (2011). Effect of Range
of Motion on Muscle Strength and Thickness. Journal of strength and
conditioning research / National Strength & Conditioning Association. 26.
2140-5. 10.1519/JSC.0b013e31823a3b15.
Poliquin, C. (1988) Five steps
to increasing the effectiveness of your strength training program. Journal of
strength and conditioning association. 10: 34-39.
Prestes, J., Frollini, A. B.,
Lima, C. D., Donatto, F. F., Foschini, D., Marqueti, R. D. C., … Fleck, S. J.
(2009). Comparison Between Linear and Daily Undulating Periodized Resistance
Training to Increase Strength. Journal of Strength and Conditioning Research,
23(9), 2437–2442. doi: 10.1519/jsc.0b013e3181c03548
Qaisar, R., Bhaskaran, S.
& Remmen, H. (2016). Muscle
fiber type diversification during exercise and regeneration. Free Radical
Biology and Medicine. 98. 10.1016/j.freeradbiomed.2016.03.025.
Radaelli, R. J Fleck, S.,
Leite, T., Leite, R., Pinto, R., Fernandesm L. Simão, R. (2014). Dose Response
of 1, 3 and 5 Sets of Resistance Exercise on Strength, Local Muscular Endurance
and Hypertrophy. The Journal of Strength and Conditioning Research. 29.
10.1519/JSC.0000000000000758.
Ratamess, N. A., Falvo, M. J.,
Mangine, G. T., Hoffman, J. R., Faigenbaum, A. D., & Kang, J. (2007). The
effect of rest interval length on metabolic responses to the bench press
exercise. European Journal of Applied Physiology, 100(1), 1–17. doi:
10.1007/s00421-007-0394-y
Schoenfeld, B. (2010) The
mechanism of muscle hypertrophy and their application to resistance training.
Journal of strength and conditioning research, 24: 2857-2872.
Schoenfeld, BJ. Science and Development of Muscle Hypertrophy. Champaign, IL; Human Kinetics, 2016.
Schoenfeld, B, Ogborn, D.
& Krieger, J. (2017) Dose-response relationship between weekly resistance
training volume and increases in muscle mass: A systematic review and
meta-analysis, Journal of Sports Sciences, 35:11, 1073-1082, DOI:
10.1080/02640414.2016.1210197
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., Sonmez, G. T., & Alvar, B. A.
(2014). Effects of Different Volume-Equated Resistance Training Loading
Strategies on Muscular Adaptations in Well-Trained Men. Journal of Strength and
Conditioning Research, 28(10), 2909–2918. doi: 10.1519/jsc.0000000000000480
Schoenfeld, B, Grgic, J.,
Ogbord, D. & Krieger, J. (2017) Strength and hypertrophy adaptations
between low- versus high-load resistance training: A systematic review and
meta-analysis. The Journal of strength and conditioning research, Ahead of
Print DOI: 10.1519/JSC.0000000000002200
Schoenfeld, B., Contreras, B.,
Vigotsky, A., Ogborn, D., Fontana, F. & Tiryaki-Sonmez, R. (2016). Upper
body muscle activation during low-versus high-load resistance exercise in the
bench press. 24. 217-224. 10.3233/IES-160620.
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., & Tiryaki-Sonmez, G. (2015).
Influence of Resistance Training Frequency on Muscular Adaptations in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(7),
1821–1829. doi: 10.1519/jsc.0000000000000970
Schoenfeld, B. J., Peterson,
M. D., Ogborn, D., Contreras, B., & Sonmez, G. T. (2015). Effects of Low-
vs. High-Load Resistance Training on Muscle Strength and Hypertrophy in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(10), 2954–2963.
doi: 10.1519/jsc.0000000000000958
Schoenfeld, B. J., Ogborn, D.
I., & Krieger, J. W. (2015). Effect of Repetition Duration During
Resistance Training on Muscle Hypertrophy: A Systematic Review and
Meta-Analysis. Sports Medicine, 45(4), 577–585. doi: 10.1007/s40279-015-0304-0
Schoenfeld, B. J., Contreras,
B., Krieger, J., Grgic, J., Delcastillo, K., Belliard, R., & Alto, A.
(2019). Resistance Training Volume Enhances Muscle Hypertrophy but Not Strength
in Trained Men. Medicine & Science in Sports & Exercise, 51(1), 94–103.
doi: 10.1249/mss.0000000000001764
Schoenfeld, B.J. & Grgic,
J. (2020) Effects of range of motion on muscle development during resistance
training interventions: A systematic review. SAGE open medicine, 8: 1-8. https://doi.org/10.1177/2050312120901559
Schuenke, M. D., Herman, J.
R., Gliders, R. M., Hagerman, F. C., Hikida, R. S., Rana, S. R., … Staron, R.
S. (2012). Early-phase muscular adaptations in response to slow-speed versus
traditional resistance-training regimens. European Journal of Applied
Physiology, 112(10), 3585–3595. doi: 10.1007/s00421-012-2339-3
Selye, H. (1950). Stress and
the General Adaptation Syndrome. Bmj, 1(4667), 1383–1392. doi:
10.1136/bmj.1.4667.1383
Serrano, A. L., Baeza-Raja,
B., Perdiguero, E., Jardí, M., & Muñoz-Cánoves, P. (2008). Interleukin-6 Is
an Essential Regulator of Satellite Cell-Mediated Skeletal Muscle Hypertrophy.
Cell Metabolism, 7(1), 33–44. doi: 10.1016/j.cmet.2007.11.011
Simão, R., Spineti, J.,
Salles, B. F. D., Matta, T., Fernandes, L., Fleck, S. J., … Strom-Olsen, H. E.
(2012). Comparison Between Nonlinear and Linear Periodized Resistance Training.
Journal of Strength and Conditioning Research, 26(5), 1389–1395. doi:
10.1519/jsc.0b013e318231a659
Snyder, B. J., & Leech, J.
R. (2009). Voluntary Increase in Latissimus Dorsi Muscle Activity During the
Lat Pull-Down Following Expert Instruction. Journal of Strength and
Conditioning Research, 23(8), 2204–2209. doi: 10.1519/jsc.0b013e3181bb7213
Snyder, B. J., & Fry, W.
R. (2012). Effect of Verbal Instruction on Muscle Activity During the Bench
Press Exercise. Journal of Strength and Conditioning Research, 26(9), 2394–2400.
doi: 10.1519/jsc.0b013e31823f8d11
Stewart, C. & Rittweger,
J. (2006) Adaptive processes in skeletal muscle: molecular regulators and
genetic influences. Journal of musculoskeletal and neuronal interactions: 6(1): 73-86.
Stone, M., O’Bryant, H. &
Garhammer J. (1981) Hypothetical model for strength training. Journal of sports
medicine and physical fitness, 21: 342-351.
Stone, M. H., Potteiger, J.
A., Pierce, K. C., Proulx, C. M., Obryant, H. S., Johnson, R. L., & Stone,
M. E. (2000). Comparison of the Effects of Three Different Weight-Training
Programs on the One Repetition Maximum Squat. The Journal of Strength and
Conditioning Research, 14(3), 332. doi:
10.1519/1533-4287(2000)014<0332:coteot>2.0.co;2
Takarada,
Y., Takazawa, H., Sato, Y., Takebayashi, S., Tanaka, Y., & Ishii, N.
(2000). Effects of
resistance exercise combined with moderate vascular occlusion on muscular
function in humans. Journal of Applied Physiology, 88(6), 2097–2106. doi:
10.1152/jappl.2000.88.6.2097
Takarada, Y., Nakamura, Y.,
Aruga, S., Onda, T., Miyazaki, S., & Ishii, N. (2000). Rapid increase in plasma growth hormone after
low-intensity resistance exercise with vascular occlusion. Journal of Applied
Physiology, 88(1), 61–65. doi: 10.1152/jappl.2000.88.1.61
Tee, J. C., Bosch, A. N.,
& Lambert, M. I. (2007). Metabolic Consequences of Exercise-Induced Muscle
Damage. Sports Medicine, 37(10), 827–836. doi: 10.2165/00007256-200737100-00001
Terzis, G., Spengos, K., Mascher,
H., Georgiadis, G., Manta, P. &
Blomstrand, E. (2010). The degree of p70S6k and S6 phosphorylation in human
skeletal muscle in response to resistance exercise depends on the training
volume. European journal of applied physiology. 110. 835-43.
10.1007/s00421-010-1527-2.
Thomas, G., & Hall, M. N.
(1997). TOR signalling and control of cell growth. Current Opinion in Cell
Biology, 9(6), 782–787. doi: 10.1016/s0955-0674(97)80078-6
Uchiyama, S., Tsukamoto, H.,
Yoshimura, S., & Tamaki, T. (2006). Relationship between oxidative stress in muscle tissue
and weight-lifting-induced muscle damage. Pflügers Archiv - European Journal of
Physiology, 452(1), 109–116. doi: 10.1007/s00424-005-0012-y
Wernbom, M., Augustsson, J.
& Thomee, R. (2007). The Influence of Frequency, Intensity, Volume and Mode
of Strength Training on Whole Muscle Cross-Sectional Area in Humans. Sports
medicine (Auckland, N.Z.). 37. 225-64. 10.2165/00007256-200737030-00004.
Willoughby, D. (1993) The
effects of mesocycle-length weight training programs involving periodization an
partially equated volumes on upper and lower body strength. Journal of strength
and conditioning research, 7: 2-8.
Wilson, J. M., Lowery, R. P.,
Joy, J. M., Loenneke, J. P., & Naimo, M. A. (2013). Practical Blood Flow
Restriction Training Increases Acute Determinants of Hypertrophy Without
Increasing Indices of Muscle Damage. Journal of Strength and Conditioning
Research, 27(11), 3068–3075. doi: 10.1519/jsc.0b013e31828a1ffa
Wilson, J. M., Marin, P. J.,
Rhea, M. R., Wilson, S. M., Loenneke, J. P., & Anderson, J. C. (2012).
Concurrent Training. Journal of Strength and Conditioning Research, 26(8),
2293–2307. doi: 10.1519/jsc.0b013e31823a3e2d
Wulf, G. (2013). Attentional
focus and motor learning: a review of 15 years. International Review of Sport
and Exercise Psychology, 6(1), 77–104. doi: 10.1080/1750984x.2012.723728
Yang, S. Y., & Goldspink,
G. (2002). Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in
myoblast proliferation and differentiation. FEBS Letters, 522(1-3), 156–160.
doi: 10.1016/s0014-5793(02)02918-6
Zanou, N., & Gailly, P.
(2013). Skeletal muscle hypertrophy and regeneration: interplay between the
myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs)
pathways. Cellular and Molecular Life Sciences, 70(21), 4117–4130. doi:
10.1007/s00018-013-1330-4
Zanchi, N. E., & Lancha,
A. H. (2007). Mechanical stimuli of skeletal muscle: implications on
mTOR/p70s6k and protein synthesis. European Journal of Applied Physiology,
102(3), 253–263. doi: 10.1007/s00421-007-0588-3
Voimaharjoittelun ohjelmoinnin muuttujia ovat muun muassa volyymi, frekvenssi, paino, harjoitteet, lihaksen toimintatapa (isometrinen, konsentrinen, eksentrinen), lepopituudet, sarjan pituus, harjoitteiden järjestys, liikepituus (range of motion) ja intensiteetti. Näitä kaikkia voi muunnella, jotta harjoittelu pysyy nousujohteisena ja kehitys ei pysähdy. Kuitenkin näistä selvästi tärkein lihaskavun kannalta on volyymi. Jos taas halutaan kehittää maksimivoimaa niin sitten pitää myös intensiteettiä olla huomattavasti harjoittelussa (katso kuva 3).
Volyymi viittaa suoritetun harjoittelun määrään tiettynä aikana (Schoenfeld, 2016). Volyymin laskeminen tehdään yleensä seuraavalla tavalla: sarjat x toistot x kuorma. Myös muitakin tapoja on, kuten esimerkiksi lasketaan volyymiksi kaikki sarjat, jotka ovat yli 75% 1RM:stä. Volyymin tasainen nostaminen pitkällä aikavälillä on tehokas harjoittelun ohjelmointikeino lihaskasvua hakiessa. Ohjelmat, joissa on useampia sarjoja, vaikuttavat selvästi positiivisemmin solunsisäiseen signalointiin ja proteiinisynteesiin verrattuna ohjelmiin, joissa on vain yksi sarja (Terzis et al., 1995; Burd et al., 2010). Lihaskasvun kannalta useampi sarja on parempi kuin yksi sarja. Samaa ajattelua voidaan soveltaa myös pidemmällä aikavälillä. Korkeampi volyymi korreloi suurempien hypertrofisten adaptaatioiden kanssa (Krieger, 2010) (katso kuva 2).
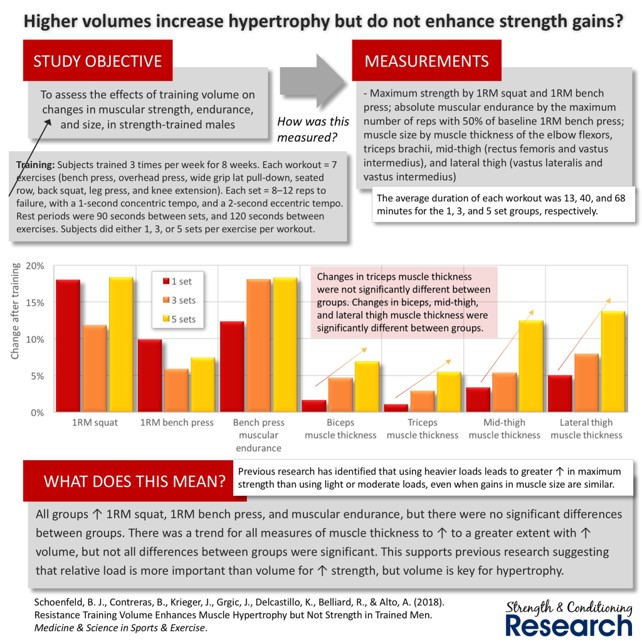
Toisaalta liian kova volyymi vähentää hypertrofisten adaptaatioiden määrää, koska rasitus on liian kova keholle. Volyymia tuleekin lisätä rauhallisesti, mutta progressiivisesti. Lisäksi periodisoinnissa tulee huomioida lepojaksot, jotta keho kykenee adaptoitumaan maksimaalisesti harjoitteluun. Radaelli ja kumppanit (2014) tutkivat 6 kuukauden ajan mikä seuraavista olisi paras lihaskasvun kannalta; 1, 3 vai 5 sarjaa. Harjoittelu toteutettiin kolme kertaa viikossa, painojen ollessa 8-12RM välillä. Tutkittavat omasivat hiukan harjoitustaustaa, eivätkä olleet siis aloittelijoita. Ne, jotka tekivät 5-sarjaa harjoittelussa, kehittyivät selvästi eniten verrattuna muihin ryhmiin. Hypertrofia jatkoi kasvamistaan viiden sarjan ryhmällä, eikä tasaantunut tällä ryhmällä niin, onkin mahdollista, että jopa vieläkin korkeampi volyymi olisi voinut kasvattaa lihaksia yhä enemmän.
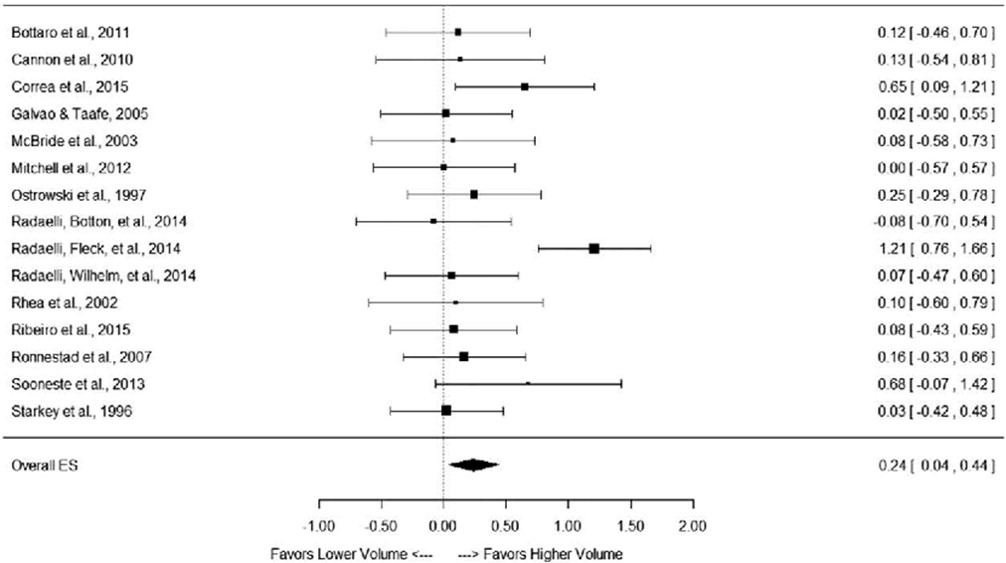
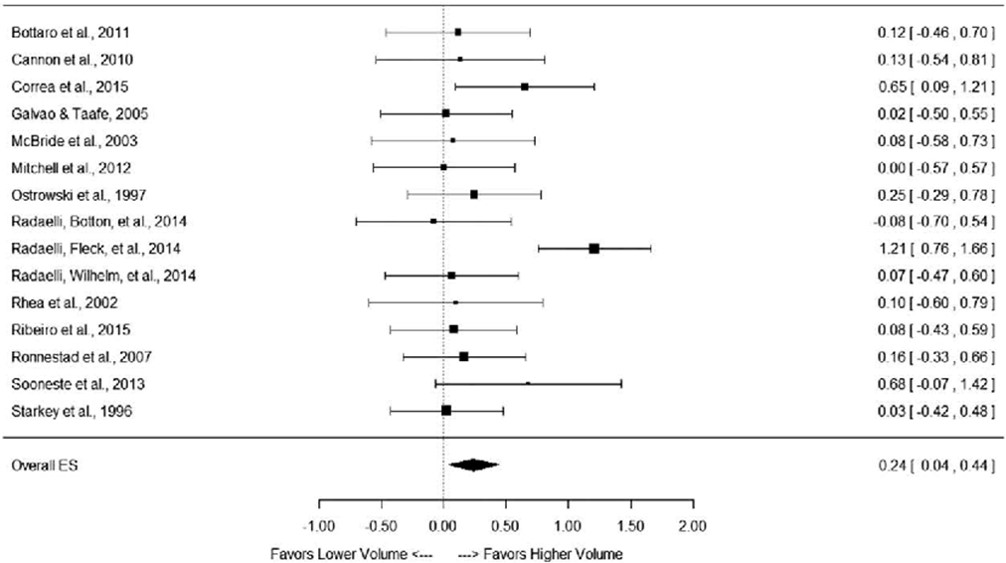
KUVA 1: Schoenfeldin ja kumppanien (2017) meta-analyysi miten harjoittelun volyymi vaikuttaa hypertrofiaan. Piirretyt neliöt kuvaavat jokaisen tutkimuksen tilastollista painoarvoa. Alhaalla on kokonaisuus, joka selvästi suosii korkeampaa volyymia.
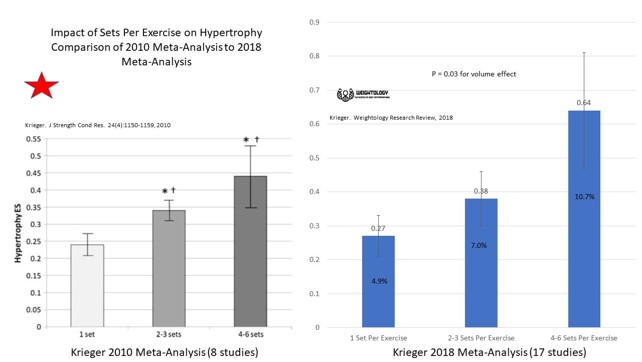
KUVA 2: Kriegerin meta-analyysien vertailu vuosilta 2010 ja 2018. Useampi sarja näyttäisi yhä edelleen olevan parempi lihaskasvulle kuin vain yksi (Krieger, 2018).
Kuinka paljon volyymia on riittävästi?
Aloittelijoille suositellaan yleensä noin 40-70 toistoa lihasryhmää kohden yhdessä harjoituksessa (Wernbom et al., 2007). Kokeneemmille nostajille taas suositus siirtyy viikkotasolle ja on noin 10-25 työsarjaa per lihasryhmä riippuen minkälaisella ohjelmoinnilla harjoitellaan. Voimapainoitteisessa harjoittelussa, jossa mennään alle 10 toiston sarjoja, voidaan puhua noin 100-150 toistosta ja bodaustyyppisessä harjoittelussa karkeasti 150-250 toistosta.
Suosittelenkin progressiivisia harjoitusohjelmia, joissa on selkeitä korkean volyymin kausia, mutta jotka sisältävät kuitenkin myös lepojaksoja. Vaikka suurempi volyymi vaikuttaakin erittäin lupaavalta, on liian suuri harjoituskuorma haitaksi kehittymiselle. Lisäksi ajatellessa vuosikymmenten mittaista ohjelmointia on hyvä huomioida kehon kestäminen. Mitä pienemmällä volyymilla pystyy kehittymään niin sen parempi. Aina on helppo lisätä volyymia ohjelmaan, mutta jos mennään koko ajan aivan maksimit piipussa niin mitä teet, kun kehitys tasaantuu? Lisää volyymia? No on muitakin vaihtoehtoja muuttujien muokkaamisessa kuin volyymi, mutta matalampi volyymi on huomattavasti nivelille ja jänteille ystävällisempää. Volyymia on aina helppo lisätä. Pitkällä aikavälillä ajatteleminen tuo parhaan mahdollisen kehityksen.
Kuva 3: Chris Beardsleyn tekemä kuva Schoenfeldin ja kumppanien (2018) tutkimuksesta, mikä selvästi osoittaa korkeamman volyymin olevan parempi hypertrofialle. Kuva on erittäin havainnollinen ja kuvastaa miten lihaksen paksuus oli korkeampi niillä, jotka tekivät enemmän sarjoja. Hypertrofian näkökulmasta volyymi näyttäisi olevan tärkein muuttuja. Toisaalta suurempi volyymi ei ollut yhteydessä suurempaan maksimivoimaan. Maksimivoima tarvitsee intensiteetin kasvua, jos sitä haluaa kehittää.
Frekvenssi
Frekvenssi tarkoittaa kuinka monta kertaa harjoitellaan tietyn ajan sisään. Esimerkiksi kuinka monta harjoituskertaa on viikossa ja kuinka monta kertaa tietty lihasryhmä harjoitellaan viikossa. Jos volyymi tasataan ryhmien välillä niin useamman kerran viikossa harjoittelevat palautuvat nopeammin, kehittyvät paremmin ja saavat enemmän voimaa sekä lihasta verrattuna niihin, jotka treenaavat vähemmän kertoja viikossa (Helms et al., 2014). Tämä perustuu siihen, että suuri volyymi yhdessä harjoituskerrassa ei ole optimaalista.
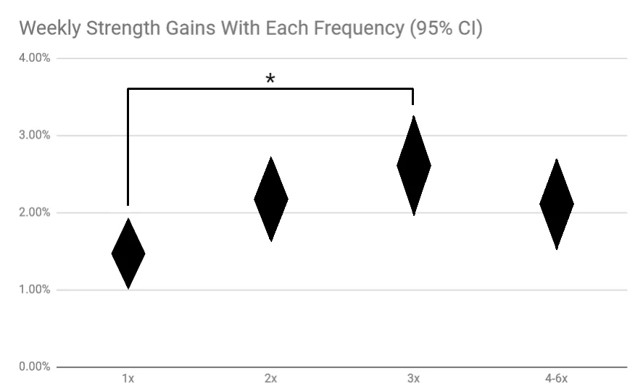
KUVA 4: Vertaillessa eri tutkimuksia voimaharjoittelun frekvenssi näyttäisi olevan optimaalisinta kolme kertaa viikossa. Yhden harjoituskerran ja kolmen harjoituskerran välillä löytyy selvä tilastollinen ero, mutta kun harjoituskertojen määriä kasvatetaan niin hyöty ei enää kasva. Toisaalta tämä tulos johtuu analysointitavasta ja eri tutkimuksia on vaikea vertailla keskenään. (Nuckols, 2018.)
Hackett:in ja kumppaneiden (2013) mielenkiintoinen kysely paljasti, että 2/3 kilpailevista mieskehonrakentajista harjoittelee tietyn lihasryhmän vain kerran viikossa ja lisäksi kukaan ei tee lihasryhmää yli kahta kertaa viikossa. Kaikki käyttivät monijaollista ohjelmaa. Monijakoinen ohjelma mahdollistaa korkeamman volyymin lihasryhmää kohden. Näissä ohjelmissa lihas viedään yleensä ihan loppuun ja silloin tarvitaan minimissään 48 - 72 h lepoa lihasryhmää kohden, ennen kuin voidaan uudestaan harjoitella sama lihasryhmä. Proteiinisynteesi kestää jopa 48 h harjoituksen jälkeen (MacDougall et al., 1995), joten voisi ajatella, että lihasryhmä pitäisi harjoitella uudestaan kahden päivän jälkeen. Proteiinisynteesi ei pysy kokonaista viikkoa ylhäällä ja jollei harjoitusta tule niin palataan jo viikossa takaisin lähtötasoon. Toisaalta jos tekee kädet maanantaina ja torstaina jo esimerkiksi selän tai rinnan niin kädet saavat epäsuorasti rasitusta ja tämä ongelma poistuu.
Kyse on myös ohjelmoinnista. Schoenfeldin ja kumppaneiden (2015) kattava tutkimus vertaili kokokehon ja jaettujen ohjelmien eroja hyvin harjoitelleilla miehillä. Koehenkilöt laitettiin pareiksi alkuvoimien mukaan ja siitä jaettiin satunnaisesti eri ryhmiin. Kahdeksan viikon jälkeen kokokehon ohjelmalla tehneet olivat saaneet enemmän hypertrofiaa yläraajoihin ja alaraajoihin verrattuna monijaollisiin ohjelmaan tehneisiin.
Miksi sitten kaikki huippukehonrakentajat tekevät monijaollisilla ohjelmilla? Nostan tässä kohtaa esiin kehon ulkopuolisen lääkityksen. Kun harjoittelua vauhditetaan kehon ulkopuolelta tulevilla hormoneilla, järkevin tapa on tehdä monijaollisilla, koska silloin proteiinisynteesi pysyy ylhäällä ja käynnissä kauemmin. Silloin voi keskittyä todella tekemään paljon volyymia lihasryhmää kohden ja todella saada voimakkaan vasteen aikaiseksi, jotta adaptaatiot lähtevät käyntiin.
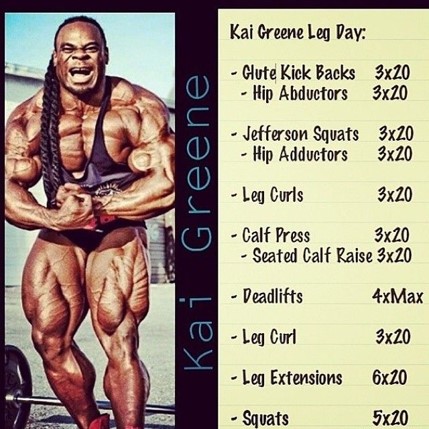
KUVA 5: Kai Greenillä tulee jalkapäivinä 700 toistoa + paljonko tekee maastavedossa toistoja. Voisi ajatella, että tallaista treeniä ei jaksa ihan kahden päivän välein vetää.
Paino
Paino voidaan laskea prosentteina maksimista, esimerkiksi 100 kilon penkillä 60 kiloa on 60%. Wernbom et al. (2007) totesivat katsauksessaan, että 6-12 on paras toistoalue hypertrofiaa varten. Tällä alueella työskentelyn ajatellaan olevan optimaalinen yhdistelmä mekaanista ja metabolista rasitusta. Painoa onkin yleisesti pidetty tärkeänä muuttujana lihaskasvun muuttujana, mutta tämä ajatus on muuttumassa. Sarjan vieminen epäonnistumiseen saakka, riippumatta painosta, johtaisi joka tapauksessa kaikkien saatavilla olevien motoristen yksiköiden aktivoimiseen (Burd et al., 2012). Toisaalta näyttäisi siltä, että maksimaalisen EMG (lihaksen aktivaation) saavuttamiseksi tulee nosta painoja, jotka ovat yli 50% (Schoenfeld et al., 2016). Pienemmillä painoilla nostelu näyttäisikin erityisesti kasvattavan tyypin 1 lihassoluja (Netreba et al., 2007). Schoenfeld ja kumppanit (2015) jakoivat yli kolme vuotta harjoitelleet miehet joko 8-12RM ryhmään tai 25-35RM ryhmään. Kahdeksan viikon jälkeen kummallakin porukalla oli kasvaneet lihakset lähes yhtä paljon ja ryhmien välille ei saatu eroja. Toisaalta 8-12 RM ryhmä oli voimakkaampi ja lihaskestävyys oli toisella porukalla paljon suurempi. Tutkimus osoitti selvästä, että myös matalammilla kuormilla voi saada aikaan hypertrofiaa, mutta käytännössä kovin moni ihminen ei jaksa vetää yksin kolmea 25-35 toiston sarjaa epäonnistumiseen saakka. Itse en olisi ainakaan selvinnyt tuosta tutkimuksesta koehenkilönä.
Vuonna 2014 Schoenfeld ja
kumppanit jakoivat 20 voimaharjoitellutta miestä joko hypertrofiaprotokollaan
tai voimaprotokollaan. Hypertrofiaprotokollassa harjoiteltiin jaetulla
ohjelmalla ja tehtiin 3 sarjaa, sekä 10 toistoa. Kun taas voimaryhmässä tehtiin
7 sarjaa, mutta vain 3 toistoa. Volyymi oli tasattu ryhmien välillä ja
tutkittavat nostivat suunnilleen yhtä paljon rautaa viikkoa kohden. Kahdeksan
viikon jälkeen hypertrofiassa ei ollut eroja ryhmien välillä, mutta
voimaryhmällä oli maksimivoima noussut enemmän. Tämä tukee volyymin tärkeyttä
hypertrofisessa harjoittelussa ja intensiteetin maksimivoimaharjoittelussa.
Tutkimuksessa oli mielenkiintoista, että voimaprotokolla kesti noin 70
minuuttia kun hypertrofinen protkolla vain 17 minuuttia harjoitusta kohden!
Kuitenkin tulokset olivat lähes samat, joten ajansäästö mielessä hypertrofinen
harjoittelu näyttäisi olevan ylivoimainen.
Schoenfeld et al. (2017)
vetivät katsauksessaan yhteen, että voima kehittyi paremmin >60% 1RM
kuormilla, mutta eroja hypertrofian välillä ei ole. Hypertrofiaa voi saavuttaa
laajalla intensiteetin skaalalla. Suosittelenkin käyttämään rohkeasti eri
painoja ja periodisoimaan voimakkaasti ohjelmaa eri intensiteeteille. Tämä
tekee hyvää myös nivelille ja jänteille. Go heavy or go home - ei ole aina
paras vaihtoehto.
Käytännön vinkkejä hypertrofisen harjoittelun suunnitteluun ja ohjelmointiin:
Variaatio on erittäin tärkeää hypertrofiassa. Lihasta tulee harjoittaa monesta eri kulmasta, liikkeen suunnasta ja asennosta. Lihaksen tiettyä osaa on mahdollista harjoitella erikseen. Lihaksen myofibriilit jaetaan eri hermolihasosastoihin, joita hermottavat eri hermon haarat. Joten jos olet kuullut, että lihasta ei kannata harjoitella monesta kulmasta, koska lihas joko aktivoituu tai ei, niin se on väärin. Motorinen yksikkö aktivoi kyllä kaikki siihen kuuluvat lihassolut, mutta esimerkiksi rectus femoris lihas (etureiden lihas) saattaa sisältää jopa yhden miljoonan verran lihassoluja, joita hermottaa 1000 liikehermosolua. Fonseca ja kumppanit (2014) vertailivat kahta ryhmää, joista toinen teki vain smith kyykkyä ja toinen ryhmä teki smith kyykkyä, jalkaprässiä, maastavetoa ja askelkyykkyjä. Volyymi oli tasattu ryhmien välillä. Variaatio, eli usempi liike, tuotti yhtenäisemmän hypertrofian kaikkiin neljään etureiden lihakseen verrattuna vain smith kyykkyyn. Lisäksi smith kyykky ei kasvattanut ollenkaan vastus medialista ja rectus femorista.
Seuraa jotenkin harjoitteluasi ja kehittymistäsi. Miten voit tietää pitääkö voluumia lisätä, jos et tiedä paljonko nostat nyt?
Ohjelmoi ja periodisoi harjoittelusi.
Lisää järkevästi ohjelmoituja korostettuja eksentrisiä harjoitteita ohjelmaan. Eksentrinen voimantuotto mahdollistaa 20-50% suuremman voimantuoton ja näin saattaa mahdollistaa suuremman mahdollisuuden myofibriilien repeämiselle ja solukalvon hajoamiselle, mikä saattaa auttaa joissain määrin hypertrofiassa. Eksentrinen harjoittelu lisää fasiculuksen (lihassolun) pituutta ja aiheuttaa hypertrofiaa niin, että myofibriilejä tulee peräkkäin. Morfologiset adapataatio eroavat hieman konsentrisen harjoittelun aiheuttamista adaptaatioista. Onkin järkevää ohjelmoida kumpiakin progressiivisesti ohjelmaan. Lisäksi eksentrisessä harjoittelussa saatetaan rekrytoida lihassoluja eri järjestyksessä kuin konsentrisessa harjoittelussa. Signalointi on eksentrisessä harjoittelussa erilaista ja erityisesti vaikuttaa MAPK aikaiseen aktivaatioon. (Allen et al., 2005; Blazevich et al., 2007.) Käytännössä lasket rauhallisesti 120-140% painoja rauhallisesti ja kontrolloidusti alas, kun taas avustaja auttaa sinua nostamaan ne konsentrisesti ylös. Isometrisestä harjoittelusta ei näyttäisi olevan nykytietämyksen mukaan hypertrofian näkökulmasta mitään hyötyä.
Nosta konsentrinen vaihe aina niin nopeasti kuin pystyt! Hidas konsentrinen vaihe ei ole optimaalista hypertrofialle ja vähentää lihaksen aktivaatiota huomattavasti (Keogh et al., 1999). Lisäksi tempon vaihtelu ei ole niin tärkeätä kuin kuvitellaan hypertrofian kannalta. Schoenfeldin ja kumppaneiden (2015) meta-analyysissä todettiin, että ei ole havaittavissa eroja hypertrofiassa yli kuuden sekunnin ja alle kuuden sekunnin toiston kestossa. Superhidas tempo (10s konsentrinen ja 4 s eksentrinen) on selvästi heikompi tapa kehittää hypertrofiaa verrattuna tavalliseen tempoon (Schuenke et al., 2012). Toisaalta toistojen tulee olla hallittuja ja elastisen energian hyödyntamistä tulee pohtia tarkasti lihaskasvuun suuntautuneessa harjoittelussa.
Harjoittele koko liikelaajuuden läpi. Koko liikelaajuuden hyödyntäminen on selvästi optimaalisempaa hypertrofialle kuin osatoistot (Pinto et al., 2012). Erityisesti etureiden kohdalla tarvitaan koko liikelaajuuden harjoittelua, muiden lihasryhmien kohdalla tarvitaan vielä lisää tutkimuksia (Schoenfeld & Grgic, 2019). Suurempi liikelaajuus näyttäisi olevan parempi, koska kova venytys ja pitkillä lihaspituuksilla harjoitteleminen aiheuttaa isomman mekaanisen jännityksen, sekä metabolisen stressin lihakseen. Toisaalta osittaisilla toistoilla on paikkansa ja aikansa suorituskykykeskeisessä, sekä myös joissain määrin hypertrofisessa harjoittelussa. Ne kuuluvat kokeneemmille harjoittelijoille ja tulee ohjelmoida järkevästi.
Matalilla kuormilla vie sarjat useimmiten konsentriseen epäonnistumiseen saakka. Muuten korkeamman kynnyksen motoriset yksiköt eivät stimuloidu. Korkeammilla kuormilla sarjoja ei tarvitse viedä loppuun saakka, koska kuorma itsessään stimuloi korkeamman kynnyksen motorisia yksiköitä. Toisaalta tämä voi välillä olla hyvä tehokeino ohjelmoinnissa. Koko ajan epäonnistumiseen saakka harjoittelu voi johtaa ylikuntoon ja IGF-1 - lepokonsentraation laskuun (Izquierdo et al., 2006), joten on äärimmäisen tärkeä ohjelmoida tarkkaan hypertrofinen loppuunviety harjoittelu.
Lihaskasvuun tähtäävässä harjoittelussa tulee miettiä lihaksen supistumista. Mielen ja lihaksen välisen yhteyden kasvattaminen auttaa lihaskasvussa parantamalla suoraan lihaksen aktivointia erityisesti harjoitellessa alle 80% painoilla (Snyder & Leech, 2009; Snyder & Fry, 2012). Jotkut salimyytit pitävät tosiaan paikkansa! Toisaalta suorituskeskeisessä harjoittelussa ulkoinen keskittymisenkohde (lopputulos) on paljon tärkeämpi ja siihen pitää keskittää huomio (Wulf, 2013).
Ravinnosta lyhyesti - älä unohda sitä! Se on hypertofisen harjoittelun tärkeimpiä asioita. Enemmän ravintoa on enemmän ravintoa, mutta liika on aina liikaa.
Tuhoaako aerobinen harjoittelu kaikki gainsit?
Jos tavoitteena on
valtavat lihakset niin onko aerobisesta harjoittelusta tällöin haittaa?
Aerobinen harjoittelu johtaa AMPK - signalointireitin aktivoitumiseen (Atherton et al.,2005). AMPK toimii energiasensorina. Sen aktivoituminen johtaa katabolisten reaktioiden käynnistämiseen, jotta saadaan lisää energiaa. Lisäksi sen aktivoituminen aiheuttaa anabolisten reaktioiden lopettamiseen. Näiden reaktioiden lopettaminen säästää energiaa, koska ne ovat energiasyöppöjä. Toisaalta anaerobinen energiantuotto johtaa Akt signalointi väylän aktivoimiseen. AMPK ja Akt toimivatkin vastapareina ja johtavat vastakkaisiin toimintoihin solussa. Tällä perusteella aerobista harjoittelua ei kannattaisi tehdä ollenkaan, mutta onko asia näin yksinkertainen?
Tämä näyttäisi olevan nykyisen tiedon valossa yksinkertaistettu versio ja aerobinen harjoittelukin voi lisätä jopa mTOR:in aktivoitumista (Benziane et al., 2008; Mascher et al., 2007). ja voimaharjoittelu AMPK väylää (Coffey et al., 2006). Aerobinen harjoittelu voi johtaa jopa lihaskasvuun. Yleensä tällaisissa tapauksissa puhutaan urheilua aloittelevista tai vanhoista ihmisistä ja lihaskasvu tapahtuu pääasiassa tyypin I - lihassoluissa (Harber et al., 2009). Harber ja kumppanit (2012) huomasivat, että tyypin I - lihassolut kasvoivat aerobisen harjoittelun seurauksena jopa 20%, mutta IIa - tyypin lihassoluissa ei ollut muutosta, paitsi ne pienenivät nuoremmilla koehenkilöillä. Nämä tulokset voivat johtua inaktiivisuuden loppumisesta ja paluusta “normaaliin” ihmisen suorituskykyyn ja kokoon. Kokeneemmilla harjoittelijoilla lihakset näyttäisivät kuihtuvan pois kovassa kestävyysharjoittelussa (Kohn et al., 2011).
Wilsonin ja kumppaneiden
(2012) meta-analyysi totesi, että yhdistetty aerobinen ja hypertrofinen
harjoittelu johti jopa 50% gainsien tippumiseen kun aerobinen harjoittelu
yhdistettiin voimaharjoitteluun. Aerobisen harjoittelun voluumilla ja
frekvenssillä on suurin vaikutus lihaskasvuun. Luultavasti, koska katabolisten
hormonien pitoisuudet nousevat ja lihasten glykogeenivarastot tyhjenevät
nopeammin (Mikkola et al., 2012).
Kun laitetaan olemassa oleva data yhteen, näyttää hyvin selvältä, että aerobisen harjoittelun ja hypertrofisen harjoittelun yhdistäminen on aina kompromissi adaptaatioissa kummankin suhteen. Mitä kokeneempi treenaaja on, sitä huonompi kombinaatio on. Aerobista harjoittelua ei kannata tehdä, jos tavoitteena on maksimaalinen lihaskasvu. Toisaalta aerobista harjoittelua kannattaa yleisen terveyden vuoksi tehdä. Kestävyysharjoittelulla on paljon erittäin positiivisia vaikutuksia terveyteen. Lisäksi kestävyysharjoittelu on (suhteellisen) hauskaa ja sitä voi harrastaa monessa eri lajissa. Suosittelen lämpimästi kestävyysharjoittelua kaikille tietyissä määrin.
Miten aerobista harjoittelua tulisi tehdä, jotta voimaharjoittelu olisi mahdollisimman optimaalista?
Intensiteetin tulee olla suhteellisen korkea. 80-85 % VO2maksimiksta tulisi olla minimi intensiteetti. Tällaisilla intensiteeteillä lepotauot suoritusten välissä tulee huomioida harjoittelussa.
Kokeile maksimaalisia sprinttejä. Nämä ovat hyvä tapa kehittää nopeutta ja saada silti hieman kestävyysharjoittelun adaptaatioita aikana. Matkaa, intensiteettiä ja palautuksia muuttamalla harjoittelun vasteista saa hyvinkin erilaisia.
Kestävyysharjoittelua tulee olla maksimissaan kolme kertaa viikossa, jos tavoitteena on maksimaalinen lihaskasvu.
Suorita, jos mahdollista, kestävyysharjoittelu ja voimaharjoittelu eri päivinä
Pyöräily näyttäisi olevan lihasten säilyvyyden kannalta parempi kestävyysurheilumuoto. Luultavasti sen takia, että iskutusta ei tapahdu ja lihasvaurioita syntyy vähemmän. Tällöin kestävyysharjoittelu ei ole niin kuluttavaa.
(Wilson et al., 2012)
Viimeisessä osassa pureudutaan harjoittelun yksilöllisiin vasteisiin ja lihaskasvuharjoittelun ohjelmointiin. Lähdeluettelo tulee viimeisen osan yhteydessä kokonaisuudessaan.
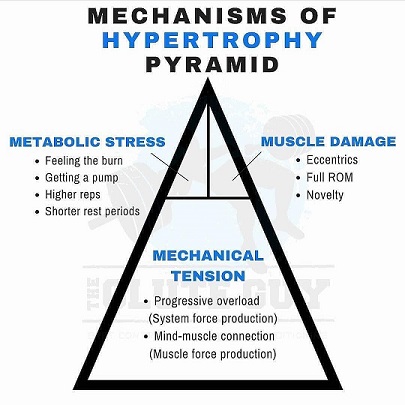
Voimaharjoittelulla voidaan saavuttaa lihaskasvua karkeasti kolmella eri tavalla. Nämä kolme eri tapaa ovat mekaaninen jännitys, metabolinen stressi ja lihasvauriot (Schoenfeld, 2010). Näistä jokainen aiheuttaa hiukan erilaisia vasteita kehossa. Mikä on tehokkain lihaskasvun kannalta?
Mekaaninen
jännitys
Tärkein tekijä hypertrofisen vasteen takana on mekaaninen jännitys. Mekaaninen kuormitus stimuloi suoraan mTOR:ia (Hornberger et al., 2006), joka stimuloi lihaskasvua. Tällä hetkellä näyttäisi siltä, että mekanosensorit (aistivat mekaanista kuormitusta) ovat herkkiä mekaanisen kuormituksen voimakkuudelle ja ajalliselle pituudelle. Nämä mekanosensorit aistivat mekaanisen jännityksen ja muuttavat stimuluksen kemiaaliseksi signaaliksi myofibriilin sisällä. Tämä johtaa signalointiprosessiin, joka lopulta muuttaa lihasproteiini synteesiin plussan, eli rakentavan puolelle.
Kuva 1: Paljolti yksinkertaistettu kaavakuva siitä, miten voimaharjoittelu aiheuttaa mekanosensoristen signalointiväylien kautta mTOR:in aktiivisuutta ja näin aiheuttan lihaskasvua. Mukaeltu: Qaisar et al., 2016.
Mekaanisen kuormituksen voimakkuus ja ajallinen pituus ovat siis tärkeimmät muuttujat lihaskasvussa. Ajallisen pituuden voi ymmärtää niin, että kuinka paljon lihas on työn alla. Se näyttäisi olevan hypertrofian kannalta tärkeä muuttuja. Lontoon kielestä poimittu time under tension (TUT) on voimaharjoittelun muuttuja, jota voidaan hyödyntää harjoittelussa. Lihaksen työmäärä ajallisesti, eli siis TUT, näyttäisi olevan yhteydessä lihaskasvun kanssa. Esimerkiksi Martineau & Gardiner (2002) stimuloivat rottien kaksoiskantalihasta ja löysivät, että TUT oli lineaarisesti yhteydessä lihaskasvun signalointiin.
Kuormituksen voimakkuuteen vaikuttaa taas lihaksen venytyksen määrä ja lihastyötapa. Erilaisia lihatyötapoja on dynaaminen, isometrinen ja eksentrinen. Näistä jokainen aiheuttaa hiukan erilaisen vasteen kehossa. Lihaskasvun osalta täydellä liikeradalla työskenteleminen näyttäisi olevan tehokkain tapa kasvattaa lihasta. Toisaalta osatoistoja ja muita erikoistekniikoita voi käyttää harjoittelussa variaationa sekä harjoittelun tehostamiseen.
Mekaaninen ylikuormitus on pitkällä aikavälillä tärkein progressiotapa lihaskasvua haettaessa. Siihen liittyy kuormituksen voimakkuuden nostaminen ja sen ajallisen pituuden kasvattaminen. Se aiheuttaa myofibriilin rakenteen hajoamisen, mikä käynnistää solun sisäisen signalointiketjun. Tämä signalointiketju johtaa lopulta anaboliseen prosessiin. Tuloksena on lihaksen koon kasvu ja supistuvien elementtien määrän nousu. Yksittäinen lihassolu kasvaa poikkipinta-alaa ja pitkällä aikavälillä tämä johtaa koko lihaksen poikkipinta-alan kasvuun.
Lihashypertrofian signalointireitit - miten lihaskasvua säädellään?
Kuva 2: Tärkeimmät solunsisäiset signalointi reitit. (Mukaeltu Schoenfeld, 2016).
PI3K / AKT - reittiä pidetään tärkeimpänä lihaskasvun säätelijänä (Jacinto & Hall, 2003; Thomas & Hall, 1997). AKT proteiinikinaasi toimii anabolisena signaloijana ja katabolisten signaalien inhiboijana. Tästä kinaasista on löydetty useita eri isoformeja, joista ainakin Akt1 on herkkä mekaaniselle intensiteetille. Tämä johti aikanaan oletukseen, että ainoastaan mekaaninen stimulus aktivoi AKT väylää, mutta näin ei kuitenkaan ole (Zanchi & Lancha, 2007). AKT aktivoi mTOR:ia ja käyttää sitä saadakseen hypertrofisia adaptaatiota aikaan. Aktivoiduttuaan mTOR vaikuttaa mRNA translaatioon kiihdyttävästi ja saa aikaan anabolisia vaikutuksia. Lisäksi sen aktiivisuus inhibitoi katabolisia reaktioita. Onkin spekuloitu, että PI3K/AKT anabolinen vaikutus tapahtuukin juuri antikatabolisten tekojen kautta (Glass, 2010). MTOR väylä voidaan kuitenkin aktivoida muitakin reittejä pitkin, kuin vain PI3K/AKT väylää pitkin, vaikka se niistä tärkein onkin. Hypertrofian signalointi onkin hyvin monimutkainen prosessi ja siihen liittyy useita eri väyliä, joita kaikkia ei vielä ymmärretä.
Kalsium riippuvaisia signalointi reittejä pidetään myös tärkeinä lihaskasvussa. Lihasaktiivisuus nostaa merkitsevästi kalsiumin tasoja lihassoluissa. Tämä muutos johtaa transkriptioon ja lopulta lisääntyneeseen proteiinisynteesiin. Kalsineuriini on näistä tärkein signalointi väylä (katso kuva 2).
AMPK toimii solun energia-anturina. Sen aktivaatio nousee, kun AMP/ATP suhde nousee. Tämä tarkoittaa, että ATP:t (kehon välittömät energian lähteet) rupeavat hupenemaan. Tämä aiheuttaa kehossa muutoksia, koska välittömästi saatavilla olevat energian lähteet ovat ensisijaisen tärkeitä lihaksessa ja niistä ei haluta luopua. Harjoittelu aiheuttaa esimerkiksi AMPK lisääntymistä. AMPK on siitä ikävä kaveri, että se vähentää anabolisia prosesseja, kuten proteiinisynteesiä ja lisää katabolisia prosesseja, kuten proteiinien hajottamista. Tämän tarkoituksena on säästää energiaa ja saada uutta energiaa käyttöön. Anaboliset prosessit ovat energiasyöppöjä, joten energian puutteen aikana ne tulee lopettaa ja katabolisia toimintoja nostaa, jotta saadaan lisää energiaa käyttöön. AMPK toimii myös mTOR:in vastavaikuttajana hidastaen näin hypertrofisia reittejä, sen aktiivisuus ei tarkoita kuitenkaan täydellistä mTOR:in hiljentymistä. Harjoittelun jälkeen mTOR ja AMPK ovat kummatkin aktiivisia. AMPK siis osallistuu myös lihashypertrofian säätelyyn. AMPK:n blokkaaminen eläimillä aiheuttaa hypertrofiaa (Goodman et al., 2011). Hypoteettisesti näyttäisikin siltä, että hyvä energiatasapaino ja ATP varastojen korkea taso vähentää AMPK väylän aktiivisuutta ja näin voisi vaikuttaa positiivisesti lihaskasvuun.
Mikä on metabolisen stressin osuus lihaskasvussa?
Metabolinen stressi on harjoitteluun liittyvää metabolien kasaantumista, erityisesti laktaatin, inorgaanisen fosfaatin ja vety-ionin. Yleensä metabolisen stressin tuntee lihaksessa pumppina, lihas turpoaa ja täyttyy verellä. Anaerobisen glykolyysin käyttäminen pääasiallisena energiantuottotapana harjoituksen aikana johtaa yleisesti metaboliseen stressiin. Karkeasti harjoituksen keston pitää olla 15 - 120 sekuntia, jotta anaerobinen glykolyysi on pääasiallisena energiantuottotapana. Lyhyet maksimivoima sarjat (>85%, alle 10s) eivät siis aiheuta metabolista stressiä, vaan kuluttavat välittömiä energianlähteitä. Koska sarja kestää pitempää ja happea ei saada niin energia tuotetaan anaerobisen glykolyysin kautta, joka tuottaa sivutuotteinaan metaboliitteja, jotka kasaantuvat. Nämä tuntuvat ”hapottavilta”, eli lihasta saattaa poltella. Lisäksi lihaksen pH tasot laskevat näiden metabolien seurauksena.
Kehonrakennustyyliseen harjoitteluun usein liitetään pitkät sarjat ja poltteen hakeminen lihakseen. Onko tämä tosiaan tehokkain tapa rakentaa lihasta? Kehonrakennustyylinen harjoittelu näyttäisikin tuottavan suuremman hypertrofisen vasteen kuin voimannostotyylinen harjoittelu (Choi et al., 1998; Masuda et al., 1999). Tämä voisi johtaa ajatukseen, että metabolinen harjoittelu on erittäin tärkeää hypertrofian kannalta. Toisaalta kun volyymi tasataan kokeilujen välillä niin tulokset eivät olekaan niin selvät (Egner et al., 2013, Schoenfeld et al., 2014). Volyymi, eli sarjat kertaa painot ovat suora mittari mekaanisen jännityksen määrästä. Näyttäisikin siltä, että metabolisen stressin tärkeyttä on lioteltu lihaskasvun osalta.
Kuva 3: Miten metabolinen harjoittelu aiheuttaa hypertrofiaa? (Mukaeltu Schoenfeld, 2016) (reactive oxygen species (ROS)) (oksidatiivinen stressi, reaktaaviset happilajit)
Miksi metabolinen stressi kuitenkin olis hyvä pitää mukana harjoittelussa, vaikka mekaaninen jännitys näyttäisi olevan tärkein tekijä lihaskasvussa?
Pitkät sarjat johtavat
metaboliittien kasaantumiseen ja väsymykseen lihaksessa. Väsytyksen
lisääntyessä ja sarjan jatkuessa myös nopeammat lihassolut joudutaan rekrytoimaan
työhön mukaan. Submaksimaalisella painolla voidaan siis myös saada kaikki
lihassolut rekrytoitua. Jos sarja viedään täydelliseen uupumukseen asti, niin kaikki
nopeatkin solut rekrytoidaan mukaan (Takarada et al, 2000; Takarada et al., 2000).
Metabolinen harjoittelu
vaikuttaa nostavasti solujen nesteytystilaan ja se aiheuttaa proteiinisynteesiä
ja proteiinien hajottamisen vähentämistä (Lang et al., 1998). Luultavasti tämä
johtuu lisääntyneen nesteen aiheuttamasta paineesta sytoskeletonia, solun
tukirankaa, ja solun kalvoa vasten. Keho kokee tämän uhkaksi solun eheyttä
kohtaan ja nesteen aiheuttama paine johtaakin ultrarakenteiden vahvistamiseen (Lang,
2007). Jo meille tutut PI3K, mTOR ja MAPK näyttäisi olevan tärkeässä roolissa
signaloimassa tätä vahvistusta. Lisäksi lihassolujen turpoaminen näyttäisi vaikuttavan
myös satelliittisolujen lisääntymiseen (Dangott et al., 2000) ja täten välillisesti
vaikuttavan lihaskasvuun.
Lisäksi metabolinen stressi on suoraan yhteydessä harjoituksen jälkeiseen kasvuhormoni tasojen akuuttiin nousuun (esim: Gordon et al., 1994; Goto et al., 2005.; Takarada et al., 2000). Laktaatin ja vetyionin kasaantuminen johtaa hormonien akuuttiin nousuun (Gordon et al., 1994). IGF-1 näyttäisi nousevan monella myös, mutta tutkimukset eivät ole tästä kaikilta osin samaa mieltä. Myös testosteronin nousu odottaa selvitystään. Metabolisen stressiä on tutkittu pääasiassa vähän harjoitelleilla ihmisillä, joten meidän tulee olla varovaisia vedettäessä johtopäätöksiä. Mekanismit metabolisen stressin taustalla voivat hyvinkin vaihdella eri ihmisillä todella paljon riippuen harjoitustaustasta.
Verenkiertoa rajoittava harjoittelu (BFR) perustuu lähes kokonaan metaboliseen stressiin. Siinä nostetaan yleisesti alle 40% painoja yhden toiston maksimista. Sen on näytetty olevan turvallista ja lisäävään anabolista signalointia, sekä lihaskasvua (Loenneke et al., 2011; Fry et al., 2010). BFR (blood flow restriction) harjoittelu tarkoittaa lihaksen verenkierron rajoittamista harjoittelun aikana. Tähän voidaan käyttää erilaisia laitteita, liinoja tai jopa polvisiteitä. Itse käytän tuollaisia pehmeitä BFR harjoitteluun tarkoitettuja siteitä.
BFR tarkoituksena on estää veren osittainen poistuminen lihaksesta. Verenkierron rajoittaminen aiheuttaa sen, että lihaksen on pakko rekrytoida nopeat anaerobiset lihassolut (eivät käytä happea). Tämä tarkoittaa nopeiden lihassolujen aktivointia jopa todella matalilla intensiteeteillä. Lisäksi veren rajoittaminen paikallisesti johtaa metaboliittien, kuten laktaattien ja vetyionien kasaantumiseen. Tämä voi johtaa suurempaan kasvuhormonin eritykseen verrattuna normaaliin reeniin. Metabolisen stressin vaikutuksesta lihaskasvuun ei olla täysin yksimielisiä, mutta jonkin verran sillä on vaikutusta. Kuitenkin tärkein hypertrofiaan vaikuttava tekijä on mekaaninen rasitus ja BFR harjoittelu ei missään nimessä korvaa normaalia harjoittelua.
Kuva 4: Käsien BFR harjoittelua.
Lisäksi BFR harjoittelu sopii erinomaisesti loukkaantuneille, jotta saadaan matalillakin intensiteeteillä aktivoitua nopeita lihassoluja. Lisäksi se on kiva lisä pumppiharjoitteluun reenin loppuun, kuten minulla ylläolevassa videossa.
Käytännön ohjeita:
- Siteen
paksuus 5-10 cm (ei liian ohuita)
- Kiristä
60-70% kireydelle siteet, jos 100% on niin kireälle kuin itse saa.
Tarkoituksena ei ole rajoittaa verenkiertoa täysin. Se on vaarallista ja
typerää.
- Käytä
20%-40% intensiteettejä ykkösmaksimista
- Käytä
siteitä reiden yläosassa tai käsien yläosassa. Ei nivelten päällä.
-
Klusterisarjat hyviä. Esim. 4x15x20% 1RM:stä. 30-60s sarjan välissä. Sen
jälkeen siteet vasta pois. Ja näitä klustereita voi tehdä 2-4. - Järkevästi
toteutettuna täysin turvallista. Käytä valmentajaa apuna, jos et tiedä mitä teet.
Pidemmät tauot – enemmän
lihaskasvua?
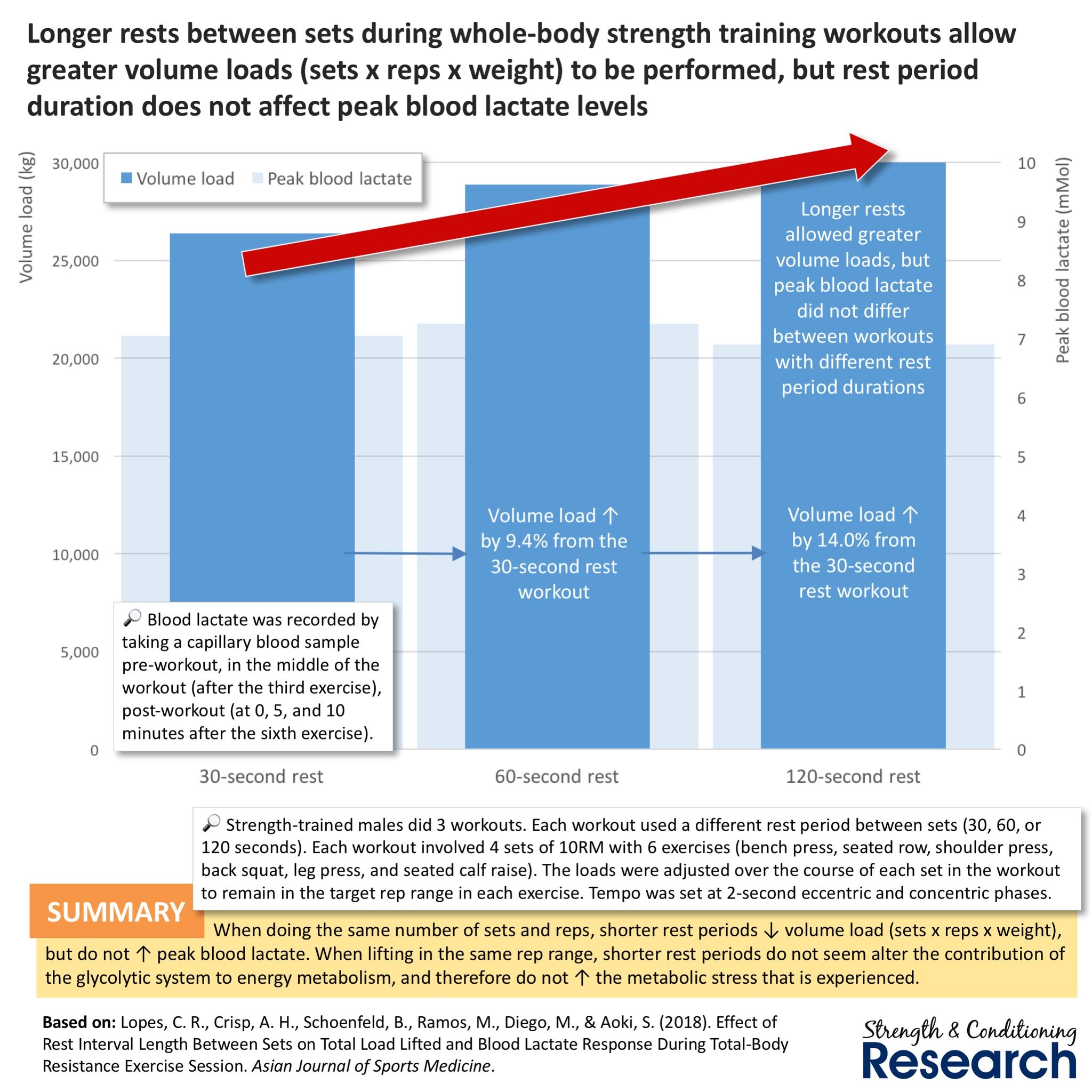
Chris Beardsley tuottama infograaffi Lopesin ja kumppanien tutkimuksesta vuodelta 2018 kuvaa hyvin sen, että aina ei kannata harjoitella lyhyillä palautusajoilla. Grafiikka kuvaa hienosti, että metabolista harjoittelua korostaessa voi kokonaisvoluumi oikeastaan jäädä pieneksi, koska lyhyet tauot rajoittavat suorituskykyä. Tämän takia on tärkeä huomioida metabolinen rasitus ilman volyymin pienentymistä. Lyhyet palautukset voivat olla sudenkuoppa. Lisäksi tutkimuksessa todettiin, että pitemmillä palautuksilla saatiin suurempi metabolinen kuorma (laktaattien kertyminen) aikaan. Tämä johtui luultavasti siitä, että pidemmillä palautuksilla voitiin tehdä suurempi määrä työtä ja saatiin kropasta enemmän irti. Alle 30 s tauoilla on todettu voluumin vähenevän yli 50% ja näin korkeampi metabolinen rasitus saadaan aikaan vähentyneellä mekaanisella jännityksellä (Ratames et al., 2007). Tämä onkin erittäin tärkeä huomioida ohjelmoidessa ja rakentaessa progressiivisia ohjelmia. Aina lyhyemmät palautukset eivät ole merkki kehityksestä.
Ahtiaisen ja kumppaneiden (2005) tutkimus on paras lepotauoista oleva tutkimus tähän mennessä. Siinä vertailtiin 2 minuutin ja 5 minuutin lepotaukojen eroja volyymi tasattujen ryhmien välillä, eli ryhmät tekivät yhtä paljon työtä. Lepotaukojen välille ei löytynyt eroja. Ainoa ongelma tässä tutkimuksessa oli, että 2 minuuttia on liian pitkä aika metaboliselle rasitukselle. Olisi ollut erittäin mielenkiintoista nähdä joko 60 sekunnin tai 90 sekunnin lepotauoilla sama tutkimus. Kaikissa muissa aiheeseen liittyvistä tutkimuksissa on mielestäni niin pahoja metodologisia ongelmia, että niistä ei kannata etsiä ratkaisua. Ohjelmoinnissa tuleekin hyödyntää sekä lyhyitä 60-90s lepotaukoja, että pitkiä lepotaukoja. Pitkillä lepotauoilla mahdollistetaan korkea volyymi ja voimatasojen palautuminen, kun taas lyhyillä lepotauoilla saadaan suurempi metabolinen rasitus aikaan. PS. Jos haluaa nähdä erittäin kivoja infotauluja voimaharjoitteluun liittyen, niin suosittelen seuraamaan Chris Beardsleytä tai Strength & Conditioning reserch:in somekanavia.
Metabolisen stressin tärkeydestä ei olla vielä täysin selvillä, koska ei tiedetä johtuvatko muutokset mekaanisesta rasituksesta vai metabolisesta rasituksesta. Näiden kahden erottaminen toisistaan on lähes mahdotonta. Kaiken lisäksi yleensä nämä kaikki tapahtuvat lihasvaurioiden kanssa samaan aikaan, joten tutkiminen on hankalaa, ellei mahdotonta. BFR harjoittelussa voidaan tehdä maltillisilla kuormilla ja saada silti aikaan metaboliittien kasaantumista. Tätä sovellettaessa tutkimuksiin ollaan huomattu, että hypertrofiaa tapahtuu myös erillään lihasvaurioista (Loenneke et al., 2014).
Lihasvauriot
Lihasvauriot voivat olla erilaisia, kuten kudosten makromolekyylien vaurioista aina sarkolemman (lihassolun solukalvon) suuriin repeämiin saakka. Lihastyön tapa, intensiteetti ja harjoituksen kesto vaikuttavat vaurioiden kokoon (Malm, 2001). Kova harjoittelu aiheuttaa vahinkoa z-levyyn ja solun tukirankaan, sekä solun ulkoiseen kalvoon. Tärkein näistä lihasvaurioista taitaa olla z-levyn repeäminen. Sarkomeeri on kahden z-levyn välinen alue. Z-levyn repeäminen tapahtuu kun myofibriilin koko kasvaa ja lateraalinen paine kohoaa. Tämä paine aiheuttaa Z-levyn repeämisen ja tämä taas seuraavan Z-levyn hajoamisen. Tämä jatkuu kunnes koko myofibriili on jakautunut pituussuunnassa. (Folland & Williams, 2007.)
Kuva 5: Z-levy näkyy kuvassa sarkomeerin reunoissa.
Lihasvauriot saavat aikaan korkeamman satelliittisolujen aktivaation. Satelliittisolut aktivoituvat, kun lihaksessa tapahtuu vauriota. Vaurioituneet lihassolun tulee nopeasti saada uusi tuma korjatakseen kudosten vauriot. Satelliittisolut auttavat tässä lisääntyen ja sulautumalla vaurioituneisiin lihassoluihin. (Dhawan & Rando, 2005.) Tulehduskipulääkkeiden ottamien saattaa estää satelliitti soluja toimimasta kunnolla ja täten vähentävän hypertrofiaa (Bondersen et al. 2004, Bondersen et al. 2006). Toisaalta mekaaninenkin stimulus ilman lihas vaurioita saa aikaan satelliitti solujen aktivoitumisen, joten asia ei ole ihan niin mustavalkoinen (Nguyen & Tidball, 2003). Mutta joka tapauksessa lihaskipuihin ei kannata ottaa buranaa, jos haluaa saavuttaa lihaskasvua.
Eksentrinen harjoittelu näyttäisi aiheuttavan kaikkein suurimmat lihasvauriot (Clarkson et al., 1986). Eksentristä lihastyötä tapahtuu kun lihasjännekompleksin pituus kasvaa. Korkeampi harjoittelun volyymi on myös yleisesti yhteydessä korkeampaan määrään lihasvaurioita (Nosaka et al., 2003). Lihasvauriot vähenevät kun samaa harjoittelua toistetaan uudestaan. Itsestään selvä ilmiö tunnetaan nimellä repeated bout effect (McHugh, 2003). Tämä johtuu siitä, että kroppa adaptoituu liikkeeseen ja vahvistaa tukikudoksia, parantaa motoristen yksiköiden synkronointia, keho oppii jakamaan tasaisemmin työn eri lihassoluille, sekä lihas synergistit toimivat tehokkaammin (Brentano & Martins Kruel, 2011). Harjoittelua tulee siis progressioida ja varioida jatkuvasti, jotta kehitys pysyy tasaisena.
Lihasvauriot heikentävät vomantuottoa, lisäävät jäykkyyttä ja turvotusta, sekä aiheuttavat lihaskipuja (DOMS:eja). Kovien lihasvaurioiden aikana harjoittelu pitää olla tarkasti mietittyä, koska jo submaksimaalinen harjoittelu nostaa sydämen sykettä ja laktaattien tuotantoa korkeammalle kuin normaalisti (Tee et al., 2007). Vaikka suorituskyky kärsii DOMS:ien aikana moni tutkija on sitä mieltä, että niiden aikaansaama tulehdustila ja proteiinien vaihtuvuus on äärettömän tärkeitä lihaskasvulle (Evans & Cannon, 1991; Finkenzeller et al., 1994). Tämähän kuulostaa hyvin järkevältä. Kroppa haluaa suojella lihasta vaurioilta ja vahvistaa sitä tulevilta vaurioilta, mutta asia ei ole kuitenkaan jälleen niin yksinkertainen. Suoraa linkkiä lihasvaurioiden ja lihaskasvun välillä ei ole pystytty osoittamaan. Lisäksi voimakkaat lihasvauriot saattavat jopa inhibitoida lihaskasvua. Vielä ei tiedetä paljonko on sopiva määrä lihasvaurioita maksimaalisen hypertrofisen vasteen aikaansaamiseksi, eikä miten lihasvauriot vaikuttavat eri harjoitustaustan omaavilla henkilöillä. Hypertrofiaa voi saada aikaan myös ilman merkittäviä lihasvaurioita (Wilson et al., 2013), mutta maksimaalisen hypertrofian aikaansaamiseksi voi olla tärkeätä saada lihasvaurioita aikaan. Toisaalta niiden optimaalista määrää ei osata vielä kertoa ja erityisesti kokeneilla harjoittelijoilla lihasvauriot voivat olla tärkeässä roolissa.
Keho vastaa lihasvaurioihin kuten tulehdukseen. Neurotrofiinit, eli hermoston kasvutekijät siirtyvät tulehduksen alueelle ja vapauttavat erilaisia substantteja hoitamaan aluetta. Neurotrofiinit saattavat signaloida muita tulehdustekijöitä lihaksen uudelleen muokkausta varten ja tätä kautta hypertrofiaa, kuten reaktiivisia happilajeja. Neurotrofiinit liitetään reaktiivisten happilajien tuottamiseen. Suoraa yhteyttä ei voida osoittaa, mutta näyttäisi kuitenkin siltä, että oksidatiivinen stressi ja reaktiiviset happilajit saattaisivat signaloida hypertrofista vastetta. Reaktiiviset happilajit signaloivat anaboliaa MAPK väylän kautta. Myös IGF-1 aktivoiminen saattaa olla tärkeä osa anabolisia signalointeja reaktiivisilla happilajeilla. Oksidatiivinen stressi suurentaa c-vitamiinitarvetta karkeasti 50-100%, eli virallisen saantisuosituksen (75mg/vrk) päälle se tekisi 100-150 mg/vrk. Monet käyttävät järkyttävän voimakkaita c-vitamiinilisä ja näiden käyttö voikin vähentää kovan harjoittelun jälkeistä lihaskipua ja nopeutta lihasten toimintakykyä (tutkimuksissa käytetty 1g). Tämä kuulostaa hyvältä, mutta tarkoittaa pienentynyttä harjoitusvastetta ja hitaampaa kehitystä. (Takarada et al., 2000, Uchiyama et al., 2006; Handayaningsih et al., 2011). Eli harjoitusvaste saadaan pienennettyä C-vitamiinilisällä, joka tarkoittaa nopeampaa palautumista, mutta toisaalta pienempää harjoitusvastetta. Tämä siis ”poistaa” osan tehdystä harjoituksesta, jonka takia palautuu nopeampaa. Eli todella suurissa c-vitaminilisien käytössä ei ole järkeä, jos tavoitteena on lihaskasvu.
Yhteenveto
Mekaaninen jännitys on tärkein muuttuja lihaskasvussa
Metabolisella stressillä ja lihasvaurioilla on roolinsa lihaskasvussa
Yhdistä harjoitusohjelmiin kaikkia kolmea
Palautusaikoja ei pidä lyhentää volyymin kustannuksella
Ensi osassa sukelletaan enemmän käytäntöön kun käsitellään eri voimaharjoittelun muuttujia lihaskasvuun tähtäävässä harjoittelussa. Käsittelyssä on mm. voluumi, frekvenssi, paino, harjoitteet, lihaksen toimintatapa, harjoitteiden järjestys ja intensiteetti. Laitan lähdeluettelon viimeisen osan yhteydessä kokonaisuudessaan.
Lihashypertrofia, eli lihassolun koon kasvu, on yksi kirjallisuuden lempiaiheitani. Omassa harjoittelussani tykkään leikkiä enemmän suorituskyvyn kanssa kuin lihaskasvun kanssa, mutta graduni käsitteli joiltakin määrin hypertrofiaa ja erilaiset artikkelit asiaan liittyen ovatkin tulleet hyvin tutuksi. Yksi erittäin mainitsemisen arvoinen kirja lihaskasvuun liittyen on Schoenfeldin Science and development of muscle hypertrophy (2016). Inspiroiduin kovasti hänen teoksestaan ja teksti lainaakin suurelta osin ajatuksiaan sieltä. Lukekaa kirja, jos haluatte kootun, tiiviin paketin aiheesta, kivoilla kuvilla. (PS. kirjassa on noin 150 sivua tekstiä, joten sen jopa jaksaa lukea läpi).
Mikä ihmeen hypertrofia?
Lihashypertrofia voidaan määritellä lihaskudoksen kasvuksi. Lihaksen supistuvien osien hypertrofia tapahtuu lisäämällä sarkomeerejä peräkkäin tai rinnakkain (Katso kuva: Sarkomeeri on lihaksen pienin toiminnallinen osa ja sisältää supistuvat aktiini- ja myosiinifilamentit).
Myofibriili, eli lihassäie, joka muodostuu peräkkäin olevista sarkomeereistä.
Kun rautaa pumppaa tarpeeksi kauan niin tapahtuu lihaksen kasvua juuri sarkomeereissä. Kaikkien yleisintä on rinnakkain tapahtuva sarkomeerien lisäys. Sarkomeerejä tulee lisää, joten lihas paksuuntuu ja kasvaa. Harjoituksen laatu vaikuttaa sihen, että minkälaista hypertrofiaa tapahtuu. Esimerkiksi eksentrinen harjoittelu vaikuttaa lihassyykimppujen (fasciculuksien) pituuteen. Toistuva kova eksentrinen harjoittelu voi siis johtaa suurempaan sarkomeerien peräkkäiseen hypertrofiaan (Lynn & Morgan, 1994). Peräkkäistä hypertrofiaa voi tapahtua myös konsentrisessa harjoittelussa, jos käytetään maksimaalista nopeutta nostovaiheessa (Alegre et al., 2006).
Lihassyykimppu on nippu lihassoluja, joita ympäröi kalvo nimeltään perimysium.
Sarkoplasmaattinen
hypertrofia
Voimaharjoittelu voi vaikuttaa myös ei-supistuviin elementteihin, eli muihinkin kuin lihakseen. Tällaisia ovat esimerkiksi kollageeni ja solunsisäinen nesteen määrä. Ilmiö tunnetaan nimellä sarkoplasmaattinen hypertrofia. Tällöin henkilö saattaa näyttää lihaksikkaammalta, mutta samanaikaisesti voimatasot eivät nouse, koska sarkoplasmaattinen hypertrofia ei vaikuta supistuviin elementteihin, jotka tuottavat voimaa.
Mistä tämä johtuu?
Glukoosia, eli
hiilihydraattia, varastoidaan lihakseen ja maksaan glykogeeninä. Glykogeeni
sitoo yhtä grammaa kohden 3 grammaa vettä. Mitä enemmän vettä on lihaksessa,
sitä isommalta se näyttää. MacDougall ja kumppanit huomasivat jo vuonna 1977,
että viiden kuukauden harjoittelun jälkeen lihaksen glykogeenivarastot nousevat
noin 66% verrattuna lähtötilanteeseen. Glykogeeni varastojen nosto saattaa olla
juuri ominaista kehonrakennustyyppiselle harjoittelulle, jossa
pääenergianlähteenä on hiilihydraatti ja energiantuottotapana glykolyysi. Harjoituksellisesti
sarkoplasmaattiseen hypertrofiaan liitetään kevyt kuorma ja korkeampi määrä
toistoja. Kun harjoitellaan lihasta väsyttävästi, keho valmistautuu väsymykseen
ja suurentaa glykogeeni varastojaan, kun taas painonnosto- ja
voimannostoharjoittelu perustuu suurelta osin välittömien energianlähteiden
käyttöön ja glykogeenivarastojen nostoa ei tarvita samassa määrin.
Tällä hypoteesilla voidaan osin perustella kehonrakentajien isommat lihakset, mutta hiukan heikommat voimatulokset, verrattuna esimerkiksi voimannostajiin ja painonnostajiin. Toisaalta ei ole mitään näyttöä siitä, että sarkoplasmaattista hypertrofiaa tapahtuisi ilman myofibriilien kasvua. Joten tässä saatettaan oikaista mutkia. Aihe on erittäin mielenkiintoinen ja odotamme lisää tutkimusta aiheen parista.
Miksi kehonrakentajat voivat pitää pitkän tauon harjoittelusta kisojen jälkeen?
Satelliittisolut ja lihaskasvu:
Satelliittisolut eivät nimestään huolimatta liity avaruuteen. Ne ovat soluja, jotka vastaavat lihassolun uudistumisesta ja huoltamisesta. Lihastuma voi vain vastata tietystä määrästä ja osasta sytoplasmaa (solukalvon sisällä oleva solulima). Tyypin II lihassoluilla tuo alue on noin kaksi kertaa suurempi kuin tyypin I lihassoluilla. Lihassolu tarvitsee siis lihastumia, jotta harjoituksen jälkeen rikottu lihas pystyy korjaamaan ja vahvistamaan itseään. Lihaskasvu tapahtuu nimenomaan tässä korjaus ja vahvistus vaiheessa, eli levon aikana. Lihas kasvaa levossa on hyvä kansanviisaus, mutta lihas tarvitsee myös ärsykkeen ennen lepoa, joten pelkkä lepo ei riitä.
Voimaharjoittelu johtaa satelliittisolujen jakautumiseen ja nopeaan lisääntymiseen. Jotkut uusista satelliittisoluista erikoistuvat lihastumiksi, luoden näin uusia lihastumia. Tämä edesauttaa nopeampaa lihassolun uudelleenrakentamista. Tätä nopeaa lisääntymistä ja tumien määrän kasvua pidetään elinehtona hypertrofialle. Satelliittisolut eivät kuitenkaan lisäänny, jollei niillä ole siihen syytä. Ne aktivoituvat, kun tarpeeksi suuri mekaaninen rasitus annetaan lihakselle. Satelliittisolut vaikuttavat myös lihaksen uudistamis-, korjaamis- ja kasvamissignalointireitteihin. (Cornelison & Wold, 1997; Folland & Williams, 2007).
Tiivistäen satelliittisolut luovuttavat tumansa myofibriileille mahdollistaen lihaksen kapasiteetin syntetisoida uusia supistuvia proteiineja. Tuma-solun massasuhde ei muutu, joten satelliittisolujen kyky luovuttaa tumansa on elintärkeä, jos lihas hypertrofiaa halutaan tapahtuvan pitkällä aikavälillä. On huomattu, että huippu voimannostajilla on selvästi suurempi määrä satelliittisoluja ja lihastumia kuin normaaleilla tallaajilla (Kadi et al., 1999; Kadi & Thornell, 2000). Nämä satelliittisolujen muutokset tapahtuvat nopeasti harjoituksen jälkeen, jopa neljän päivän sisällä yhdestä harjoituksesta (Moore et al., 2005).
Miksi Phil Heath ja muut huippukehonrakentajat voivat siis pitää 2-4 kokonaista kuukautta kisojen jälkeen taukoa harjoittelusta ja sitten palata erittäin nopeasti kisakuntoonsa? Syy löytyy luultavimmin satelliittisoluista. Egner et al. (2013) antoivat hiirille 14 päivän testosteronia, joka johti 77% lihashypertrofiaan ja 66% lihastumien kasvuun. Lihassolut palautuivat takaisin lähtötasolle kolmessa viikossa testosteroni kuurin jälkeen, mutta tumien määrä pysyi korkealla vielä kolmen kuukauden ajan. Satelliittisolut ovatkin solun omia muistimekanismeja, jotka auttavat anabolisissa reaktioissa ja nopeuttavat jälleen hypertrofiaa harjoittelun jatkuessa tauon jälkeen. Jo saavutetun lihasmassan takaisin saaminen on siis “helpompaa” kuin uuden rakentaminen.
Hormonit
ja lihaskasvu
Hormonit vaikuttavat
suuresti lihashypertrofiaan, mutta niistä voisi kirjoittaa ja on kirjoitettu omia
kirjoja, joten jätän ne tässä katsauksessa pieneen osaan. Mainitsen muutaman tärkeän
asian hormoneihin liittyen. Hormonien basaalikonsentraatiot (normaali taso)
vaikuttavat kasvuun ja lihaksen uudistumiskapasiteettiin suuresti (Conboy et
al., 2005). Tärkeimmät anaboliset hormonit ovat testosteroni, insulinin kaltainen
kasvutekijä IGF-1, kasvuhormoni ja insuliini. Kasvuhormonia erittyy eniten
syvässä unessa (pois lukien harjoituksen aikainen nousu). Joten muistakaahan
nukkua paljon syvää unta. Kasvuhormoni stimuloi lipolyysiä (rasvojen hajotusta)
ja vaikuttaa ravinteiden sisäänottoon soluun, sekä solujen kasvuun.
Kasvuhormonin anabolinen vaikutus tapahtuu suurimmaksi osaksi IGF-1 kautta. Kasvuhormoni
on steroidihormoni ja sitä ei varastoida solussa varastorakkuloihin, toisin
kuin peptidihormoneihin ja sen vaikutus kehoon on hiukan erilaista.
Kasvuhormonin eritys normaalissa päivärytmissä. Kova harjoittelu ja uni aiheuttavat akuutisti kovimmat nousut veren kasvuhormonitasoihin. (Mukaeltu Guyton & Hall, 2011.)
Kasvuhormonin muutama tärkein tehtävä:
Vahvistaa ja lisää luiden mineralisaatiota
Lisää lihasmassaa sarkomeerisen hypertrofian kautta
Miten steroidihormoni, eli kasvuhormoni, sitten toimii solussa? Steroidihormoni diffusoituu, eli siirtyy, solukalvon läpi ja kiinnittyy reseptoriproteiiniin. Tämä reseptori yhdessä steroidihormonin kanssa siirtyy tai siirretään tumaan ja siellä se sitoutuu DNA-juosteeseen kromosomissa. Siellä tapahtuu transkriptio, jossa RNA-polymeraasiin kopioidaan DNA:ssa oleva geneettinen koodi. Tämä geneettinen koodi on tällä kertaa se, että missä järjestyksessä aminohapot asetellaan proteiiniksi. RNA:ssa on nyt siis tieto siitä, miten proteiini tulee rakentaa. Sen eteen lisätään m-kirjain ja saadaan mRNA (messenger RNA), se diffusoituu sytoplasmaan, eli solukalvon sisällä olevaan solulimaan ja edistää translaatiota ribosomeissa. Translaatiossa mRNA:n sisältämä tieto käännetään geneettisen koodin mukaisesti aminohappojen järjestyksesi. Ribosomin pinnalla tapahtuu itse proteiinisynteesi. Eli steroidihormoni saa aikaan sen, että DNA:sta kaivetaan tieto mihin järjestykseen aminohapot pitää asettaa, jotta saadaan aikaan haluttu proteiini. Tämän jälkeen viedään viesti tästä eteenpäin ja rakennetaan itse proteiini. Steroidihormoni edistää siis translaatiota, mutta vaikutus on suhteellisen hidasta.
Taiteilijan näkemys siitä miten steroidihormoni toimii.
Kasvuhormonin eritystä stimuloivat tekijät
Kasvuhormonin eritystä inhiboivat (estävät) tekijät
Alhainen veren glukoosi
Korkea veren glukoosi
Alhainen veren rasvahappopitoisuus
Korkea veren rasvahappopitoisuus
Nälkiintyminen tai paasto, proteiinien puute
Ikääntyminen
Trauma, stressi, jännitys
Liikalihavuus
Liikunta
Kehon ulkopuolinen kasvuhormoni
Testosteroni, estrogeeni
Somatomediinit
Syvä uni
Hormonien vaikutus lihaskasvuun on suuri! Asiaa on tutkittu rotilla paljon ja tässä on vertailtu kahden rotan kasvukäyriä ajan suhteen kun toinen rotta sai päivittäin kasvuhormonia ja toinen ei. (Mukaeltu Guyton & Hall, 2011.) Insuliini on tullut ryminällä kehonrakennukseen ja syy löytyy sen tehokkuudesta. Yksin se ei ole tehokas aine, mutta yhdistettynä kasvuhormoniin se on todella tehokas. Yhdessä insuliini ja kasvuhormoni aiheuttavat suuren lisäyksen lihaskasvuun. (Mukaeltu Guyton & Hall, 2011).
Muut tärkeät hormonit
lihaskasvun kannalta
Testosteronia on miehillä
keskimäärin kymmenkertainen määrä verrattuna naisiin. Tämän uskotaan olevan syy
siihen, miksi miehillä on yleensä suurempi määrä lihasta kuin naisilla. (Harridge,
S., 2007.) Testosteroni vaikuttaa suoraan proteiinisynteesiin ja lisää muita
anabolisia hormoneita verenkierrossa, kuten kasvuhormonia ja IGF-1:sta.
Näistä anabolisista
hormoneista harjoittelulla ei ole vaikutusta Insuliiniin. Harjoittelu saattaa
jopa estää sen eritystä. Insuliiniin voi vaikuttaa erilaisilla ravitsemuksellisilla
asioilla kuten makroravinteiden määrällä ja ajoituksella.
Akuutti hormonaalinen vaste harjoitteluun. Kuva demonstroi suuremman vasteen jo voimaharjoitelleilla miehillä. Tämä luultavasti johtuu suuremmasta aktiivisesta lihasmassan määrästä, eli tulee enemmän väsymystä ja saadaan kropasta enemmän irti. Huomioitavaa on myös paljon suurempi keskihajonta, joka viittaa harjoitelleiden miesten suurempaan yksilölliseen vasteeseen. (Ahtiainen et al., 2003).
Akuutilla hormonaalisella vasteella on jonkinlainen yhteys hypertrofiaan ja aivolisäkkeen erittämän kasvuhormonin määrän kanssa. Metaboliittien kertyminen vaikuttaa akuutin hormonaalisen vasteen määrän. Erityisesti H+-ioni on toimiva kasvuhormonin akuuttia eritystä lisäävänä. Kansankielisemmin H+-ioni tunnetaan maitohappona. Esimerkiksi maksimivoimaharjoittelu tuottaa paljon pienemmän akuutin kasvuhormonivasteen verrattuna kehonrakennusharjoitteluun, koska toistomäärät ovat lyhyempiä. Testosteronin akuutista vasteista on paljon erilaisia tutkimuksia, luultavammin, koska sukupuoli, ikä ja harjoitustausta vaikuttavat niin suuresti testosteronin synteesiin ja näin syntyy ristiriitaisia tuloksia. Akuuttia hormonaalista vastetta ei voida suoraan yhdistää lihaskasvuun, mutta sillä saattaa olla jonkinlainen merkitys sen kanssa. Yksi esitetyistä vaihtoehdoista on se, että sen pääasiallinen tehtävä saattaakin olla ravinteiden mobilisointi lihaskasvua varten.
Myokiinit
Myokiinit on uusi nimi sytokiineille, jotka ovat proteiinirakenteisia solujen välisiä viestinnän välittäjäaineita. MGF:ää (Mechano growth factor) pidetään erittäin tärkeänä myokiininä lihaskasvua varten. MGF:än nousua on erityisesti havaittu hyvin lihashypertrofiaan respondoivilla ihmisillä ja se saattaakin olla tärkeä osa prosessia lihaksen uudelleen muokkauksessa ja hypertrofiassa (Bamman et al., 2007). MGF:ällä on anabolinen ja antikatabolinen vaikutus. Se stimuloi suoraan lihasproteiinisynteesiä ja inhibitoi proteiinien hajoitusta vähentämällä transkription aktiivisuutta. Lisäksi MGF vaikuttaa satelliittisoluihin aktivoitumiseen ja nopeaan lisääntymiseen, mutta ilmeisesti ei erillaistumiseen (Hill & Goldspink, 2003; Yank & Goldspink, 2002).
Interleukiinit ovat myös myokiinejä, jotka vaikuttavat harjoituksen jälkeen positiivisesti satelliitti solujen nopeaan lisääntymiseen ja satelliittisolujen tumien luovuttamiseen. Nämä vaikuttavat myös kollageenin synteesiin. (Serrano et al., 2008.) Harjoittelu nostaa akuutisti IL-6 tasoja jopa 100 kertaisiksi. Tunnetuin interleukiini taitaa olla myostatiini. Myostatiini on lihaskasvun negatiivinen säätelijä. Se vähentää myofibrillistä proteiinisynteesiä ja saattaa vaikuttaa satelliitti solujen aktivaatioon (Zanou & Gailly, 2013). Lihasmassa tuplaantuu hiirillä, joilta on häiritty myostatiinigeeniä (McPherron et al., 1997). Lukuisia muitakin myokiinejä on löydetty ja löydetään jatkuvasti. Näiden vaikutuksia tutkitaan jatkuvasti ja tulevaisuudessa olemmekin asian suhteen huomattavasti fiksumpia.
Belgian Blue - niminen karjalaji, jolta puuttuu myostatiinigeeni. Äärimmäinen hypertrofia ei ole kuitenkaan tällä lajilla johtanut mitenkään parempaan suorituskykyyn.
Ensi osassa keskitytään hypertrofian mekanismeihin. Laitan lähdeluettelon viimeisen osan yhteydessä kokonaisuudessaan.
Maksimivoima toimii pohjana monelle muulle ominaisuudelle. Voimaa tarvitaan muun muassa siirtämään urheilijan omaa kehoa, vastustajaa tai välinettä. Voimaa tarvitaan erityisesti kuulantyönnössä, jossa pitää siirtää pienessä betoniympyrässä rautapallo mahdollisimman kauas itsestä. Knoppitietona miesten kuulantyönnön ME-työntö 23,12 m on kuulaan tuotettuna voimana noin hevosvoiman luokkaa. Yksi hevosvoima vastaa suunnilleen 735,5 wattia. Kokonaisteho on luontaisesti tätä paljon suurempi, jopa kahdeksankertainen. (Yrjölä, 2000.)
Muissa
lajeissa tarvittavan voiman määrä vaihtelee lajikohtaisesti. Tähän vaikuttaa
paljolti, missä asennoissa voimaa tuotetaan, kuinka nopeasti sitä tulee
tuottaa, millaisissa liikkeissä sitä tulee tuottaa ja kuinka monta kertaa
peräkkäin. (Häkkinen & Ahtiainen, 2016.)
Voima voidaan jakaa nopeusvoimaan, maksimivoimaan ja kestovoimaan. Maksimivoimalla tarkoitetaan suurinta voimatasoa, jonka lihas tai lihasryhmä kykenee tuottamaan tahdonalaisessa kertasupistuksessa. Tässä tekstissä pureudutaan puhtaasti maksimivoimaharjoitteluun. Tiivistetysti maksimivoimaharjoittelun tarkoitus on kehittää hermolihasjärjestelmän kykyä tuottaa voimaa ja tukikudosten rasituskestävyyttä. (Häkkinen & Ahtiainen, 2016.)
Kuulantyöntö ja
maksimivoimaharjoittelu
Kuulantyöntö on loistava esimerkki maksimivoiman merkityksestä lajissa. Kuulantyöntäjä tarvitsee todella kovat maksimivoimatasot, jotta naisilla 4-kiloinen ja miehillä 7,26-kiloinen kuula liikahtaa ringistä mahdollisimman pitkälle. Voimantuottoaika kuulantyönnössä on vain noin 1,3 - 1,5 sekuntia, ja itse työntövaihe kestää siitä 0,2 - 0,25 sekuntia (Yrjölä, 2000). Voima pitää siis kyetä myös tuottamaan nopeasti. Suuresta määrästä voimaa ei ole hyötyä, jos sitä ei pysty tuottamaan räjähtävästi rautapalloon. Kuulantyöntökilpailun aikana voimaa tuotetaan vain kuusi kertaa, ja jokainen suoritus on asyklinen eli tapahtuu vain kerran. Kuulantyöntö on siksi loistava tarkastelunkohde, koska maksimivoimatasot tulee olla äärimmäisen korkealla, ja voima tulee tuottaa nopeasti.
Verrattuna vaikkapa voimanostajiin kuulantyöntäjän tulee olla paljon räjähtävämpi ja monipuolisempi urheilija. Voimannostajan ei tarvitse nostaa painoa nopeasti, kunhan paino vain nousee. Kuulantyöntäjän taas pitää lyhyessä ajassa tuottaa voimaan kuulaa, mutta toisaalta kuulantyöntäjien ei tarvitse periaattessa olla maksimivoimatasoiltaan yhtä voimakkaita kuin voimannostajien. Kuulantyönnön voimantuottoaika on hyvin lyhyt. Se on vain 0,2 – 0,25 sekuntia kun tahdonalaisen maksimaalisen voimatason saavuttaminen vie jopa 0,5 – 2,5 sekuntia riippuen lihaksesta, liikkeestä, harjoitustaustasta ja geeneistä (Mero, Ahtiainen & Häkkinen, 2007). Tämän takia voimanostajien harjoittelu on hieman erilaista. Painonnostoharjoittelu näyttäisi sopivan erityisen hyvin kuulantyöntäjille. Se johtuu painonnostoliikkeiden räjähtävämmästä luonteesta verrattuna voimanostoon.
No kuinka vahva
kuulantyöntäjän tulee sitten olla?
Tarvittavia
voimatasoja on yrittänyt lisäkseni määritellä Auvinen (2011) (Räntilä, 2015)
(taulukot 1 ja 2.). Ylhäältä voi lukea halutun tuloksen ja siihen vaadittavat
voimatasot. Taulukko on suuntaa antava, ja sitä voi käyttää apuna urheilijan
harjoittelun ohjelmoinnissa. Urheilijan yksilölliset vahvuudet ja heikkoudet
kannattaa huomioida tarkkaan.
Kaikki
ominaisuudet heittolajeissa ovat rakennettavissa, ja melkein jokaisesta
innostuneesta heittäjästä olisi mahdollisuus saada hyvän tason heittäjä
Suomessa. Ominaisuuksien tasainen rakentaminen on kuitenkin valmennuksellisesti
vaikeaa, ja se vaatii pitkäjänteistä työtä vuodesta toiseen. Fyysiset
ominaisuudet eivät kehity hetkessä tai nopeasti vaan tasaisesti ja hitaasti.
Ominaisuusnormistot (taulukot 1 ja 2) ovat oiva väline nuorelle heittäjälle
seuraamaan omaa kehitystään. Omien vahvuuksien ja heikkouksien löytäminen
helpottuu taulukoiden avulla. Taulukoita luetaan niin, että tulokset ovat ylärivissä
ja fyysiset ominaisuudet pystyrivissä. Esimerkiksi jos mieskuulantyöntäjä
haluaa työntää yli 20 metriä niin tulisi hänen rinnallevetonsa olla 175 kg ja
penkkipunnerruksen 215 kg.
Karoliina Lundahl, suomalainen painonnoston maailmanmestari vuosilta 1994 ja 1998, edusti Suomea Atlantan olympiakisoissa vuonna 1996 myös kuulantyönnössä. Hänen voimatasonsa olivat erittäin korkeat ja onkin spekuloitu, että hänen voimatasonsa olisivat riittäneet huomattavasti parempaankin kuulantyöntötulokseen. Lundahlin ennätys on 18,28 m. Hänen painonnostoennätyksillään (tempaus 107 kg ja työntö 127 kg) olisi Auvisen taulukon mukaan pitänyt työntää 20 metriä (katso taulukot 1 ja 2).
Nykytyöntäjistä esimerkiksi Suvi Helin on yhdistänyt painonnoston ja kuulantyönnön huipputasolla. Hänen ennätyksensä on 15,96 m. Suvi Helinin ennätykset vuonna 2015 olivat tempauksessa 85 kg ja työnnössä 105 kg. Näillä hänen olisi pitänyt työntää yli 17 metriä, mutta hän on työntänyt “vain” 15,96 m. Miksi nämä naiset eivät ole työntäneet pidemmälle hurjista voimatasoista huolimatta? Selitys löytyy yksinkertaisesti tekniikasta. Voima tulee tuottaa välineeseen lyhyessä ajassa vaativien liikesarjojen kautta. Nyt voimaa ei ole saatu tuotettua välineeseen. Tämän takia kuulantyöntäjä viettää harjoitusajasta karkeasti vain puolet salilla, joissakin tapauksissa jopa huomattavasti vähemmänkin (Zatsiorsky & Kramer, 2006).
Painonnoston ja kuulantyönnön yhdistäminen huipputasolla on siis erittäin haastavaa. Lajit vaativat hyvin erilaista erikoistumista harjoittelun ja kehon jänteiden sekä nivelten adaptaatioiden suhteen. Harjoittelu kuulantyönnön huipputasolla vaatii hyvin paljon työntämistä ja erilaisia drillejä sekä rotaatiovoiman kehittämistä. Painnonostoharjoittelu vaatii kasvavan harjoittelumäärän omalta osaltaan. Näiden yhdistäminen on ajankäytöllisesti ja kehon jaksamisen takia hyvin vaikeaa. Toisaalta nuorille urheilijoille näiden lajien yhdistäminen sopii erinomaisesti.
Taulukkoa 1 varten on kerätty aineistoa suomalaisista huipuista ja sen takia esimerkiksi miesten kuulantyönnössä pituus ei muutu. Suomalaisessa kuulantyönnössä on vaikuttanut paljon pyörähtäjiä ja heidän antropometriansa ei ole yhteydessä kuulantyöntö tuloksen kanssa. Toisin kuin pakittajilla. Jos pakittajia olisi aineistossa enemmän, olisi pituus varmasti merkitsevämmässä roolissa. Lisäksi Suomessa on aika vähän huippunaistyöntäjiä ollut, joten naisten taulukon aineisto on selvästi painottunut 14 metrin ja 17 metrin välille. Uskon itse, että yli 17 metrin työnnöt vaativat hiukan kovemmat maksimivoimatasot kuin mitä taulukossa nyt on.
Kuten kaikkien voimaharjoittelijoiden kuulantyöntäjänkin tulee käyttää 2-3 vuotta tekniikoiden hiomiseen, ennen kuin nousujohteinen voimaharjoittelu voi alkaa. Tätä tekniikkaharjoittelua on helppoa tehdä 11 - 15-vuotiaana, jolloin harjoituksiin voi yhdistää liikkuvuuden, kestovoiman ja nopeusvoiman elementtejä. Vanhemmalla iällä, kun voimaa jo on, tekniikoiden rakentaminen alusta on yleensä varsin mälsää puuhaa. Kuulantyöntäjälle parhaat liikkeet riippuen työntäjän antropometriasta ovat tempausvariaatiot, rinnallevetovariaatiot, työntövariaatiot, kyykkyvariaatiot, maastavetovariaatiot ja erilaiset apuliikkeet. Kuulantyönnössä korostuu jalkojen vertikaalinen voimantuotto ja yläkropan horisontaalinen voimantuotto, jonka kehittämiseen esimerkiksi penkkipunnerrus on hyvä liike.
Harjoittelun yksi tärkeimpiä tekijöitä on nousujohteisuus. Harjoitusmäärien tasainen nostaminen voimaharjoittelussa tulee tehdä tasaisesti monen vuoden aikana. Suuria voimaominaisuuksia ei saavuteta nopeasti tai lyhyen aikavälin runsailla volyymilisäyksillä. Lisäksi äkkinäiset volyymilisäykset altistavat urheilijan turhalle loukkaantumisriskille.
Volyymi saadaan kertomalla toistot
sarjalla ja painolla, esimerkiksi 10 sarjaa 10 toistoa 100 kilolla tekee
yhteensä 10 000 kiloa eli 10 tonnia. Yleensä volyymiin lasketaan vain ne sarjat,
jotka tehdään yli 70 % intensiteetillä lasketusta ykkösmaksimista. Kuulantyöntäjän tulee ohjeellisesti nostaa
minimissään 900 tonnia vuodessa ja maksimissaan 1450 tonnia vuodessa.
Valmistavalla kaudella ohjeellisesti tonnit vaihtelevat 24 - 40 välillä yhdellä
harjoitusviikolla ja kilpailukaudella 8 - 12 tonnin välillä. (Bompa & Haff, 2009)
Harjoittelun kehitystä voi seurata myös laskemalla tehdyt sarjat, jotka ylittävät tietyn tuntemuksen rajan (esim. RPE taulukko) tai sarjat, joissa nopeus tippui tietyn prosenttiosuuden verran (esim. 20 % maksiminopeudesta, tämä vaatii tosin nopeuden mittaamisen).
TAULUKKO 3: Kuulantyöntäjän klassinen ja paljon käytetty harjoitusvuoden jaottelu. Vaikka painopisteet on jaoteltu, tulee voimaharjoittelu ja työntäminen pitää mukana kaikilla harjoittelukausilla. (Mukailtu Yrjölä, 2000). Nykyään suositaan myös paljon blokkiperiodisaatiota klassisen periodisaation rinnalla.
Hypertrofisella harjoittelulla tarkoitetaan lihassolun poikkipinta-alan kasvuun tähtäävää harjoittelua. Harjoittelujakson tavoitteena voi olla rasvattoman massan lisääminen, kudoksien vahvistaminen, mahdollisesti myös rasvamassan vähentäminen ja työkapasiteetin nostaminen, Tämä luo pohjat intensiteetin ja volyymin nostolle. Lisäksi luodaan hermolihasjärjestelmään tasapaino ja ehkäistään puolieroja ja vaikutetaan loukkaantumisten ehkäisyyn. (Bompa & Haff, 2009.)
Hypertrofiaa voidaan saavuttaa laajalla skaalalla erilaisia harjoitusohjelmia. Tärkeimmät vaikuttavat muuttujat harjoitusohjelmat lopputulokseen ovat kuorma, intensiteetti, nopeus, volyymi, sarjojen välinen lepo, liiketyyppi ja aktiivisen lihasmassan määrä. Volyymin ajatellaan olevan erittäin tärkeä tekijä morfologisten adaptaatioiden saavuttamisiksi (Krieger, 2010). Volyymin nosto harjoitusohjelmassa on helppo progressiotapa hypertrofisen jakson aikana. Yksi lihaskasvun tärkeimmistä ärsykkeistä on mekaaninen kuormitus (Schiaffino et al., 2013). Myös metabolisella rasituksella ajatellaan olevan rooli hypertrofiassa (Schoenfeld, 2013) ja myös jossain määrin voimassa (Rooney et al., 1994; Schott et al., 1995). Näiden kummankin yhdistäminen voi olla optimaalista anatomisten adaptaatioiden saavuttamiseksi.
KUVA 1: Bret Contreras kuvasi hyvin mekaanisen ja metabolisen rasituksen sekä lihasvaurioiden suhdetta. Hypertrofisessa harjoittelussa mekaaninen jännitys ja progressiivinen kuormitus ovat tärkeintä. Metabolisella kuormituksella ja lihasvauriolla on osuus kokonaisuudessa, mutta se on pienehkö.
Maksimivoimaharjoittelulla kehitetään sitä maksimaalista voimaa, mikä kyetään tuottamaan. Tällöin yleensä harjoittelu keskittyy hermostollisen ja lihakseen liittyvän kapasiteetin maksimaalisten adaptaatioiden saavuttamiseen. Kehittyäkseen maksimivoima tarvitsee korkean intensiteetin.
Maksimaalisella yrittämisellä (aggressiivinen nopeusvoimaperiaate) rekrytoidaan mahdollisimman täydellisesti nopeat motoriset yksiköt eli nopeat liikehermot ja niiden päissä olevat nopeasti supistuvat lihassolut. Tämä on erittäin tärkeää maksimivoimajaksolla kuin myös sitä seuraavalla nopeusvoimajaksolla.
TAULUKKO 4: Miten voimaharjoittelu voitaisiin käytännössä ohjelmoida.
Lisää
räjähtävää voimaa
Räjähtävävoiman ja pikavoima jaksolla on tarkoitus oppia tuottamaan voimaa mahdollisimman paljon lyhyessä ajassa. Ohjelmoinnista riippuen harjoittelussa voi tehdä myös yhdistettyä voima- ja nopeusharjoittelua. Kuulantyöntäjän harjoittelussa tulee muistaa vielä lajivoiman lisääminen ohjelmointiin. Lajivoiman kehittäminen tapahtuu yleensä eripainoisia välineitä työntämällä. Taulukosta 4 voi katsoa karkean esimerkin vuositason ohjelmoinnista kokeneemmalle urheilijalle.
Räjähtävä voima tarkoittaa kykyä käyttää maksimaalista voimaa mahdollisimman lyhyessä ajassa. Parhailla kuulantyöntäjillä 21 metrin työnnöissä on mitattu heittokäden käsityönnön voimaksi 50-60 kg. Näiden työntäjien maksimaalinen käsien ojennus - eli penkkipunnerrustulos - vaihtelee tyypillisesti 220-240 kilon välillä eli 110-120 kg kättä kohden. Itse lajisuorituksessa on tällöin onnistuttu tuottamaan noin 50 % maksimaalisesta käytössä olevasta voimasta. Heittolajeissa voi parantaa voimantuottoa kahdella tavalla. Ensinnäkin voi lisätä maksimaalista voimaa. Toinen tapa on lisätä voimantuottonopeutta. Heittäjän uran alussa lapsuudessa ja nuoruudessa voi olla viisasta kehittää maksimaalista voimaa ja yleisesti lihaksistoa unohtamatta nopeuden herkkyysvaiheita. Tämä antaa laajan pohjan voimantuottonopeusharjoittelulle. Toisinpäin tehtynä pelkkää nopeutta kehittäen maksimaalista voimaa ei saada enää uralla riittävästi käyttöön. Tehokkainta on näiden kummankin yhdistäminen optimaalisesti ja yksilöllisesti. (Zatsiorsky & Kramer 2006.)
On tärkeää tehdä maksimivoimajakso ennen
räjähtävän voiman jaksoa, koska tehon lisäys voi tapahtua niin maksimaalisen
voiman kuin nopeuden lisäyksenä tai molempien kasvulla. Jakson pituus voi
vaihdella yhdestä kolmeen kuukauteen, riippuen kisataanko hallikaudella.
Räjähtävän voiman/pikavoiman kausi tulee sijoittaa kilpailuun valmistavalle
kaudelle. Sen tarkoituksena on muuntaa saavutetut korkeammat voimatasot lajin
käyttöön. On kuitenkin tärkeä muistaa ylläpitää maksimivoimatasoja, koska
muuten nopeus kärsii. (Bompa & Haff, 2009.)
Siirtymävaihe on kuulantyöntäjälle hyvin tärkeä. Tämä jakso ajoittuu heti kilpailukauden jälkeen ennen uuden kauden aloittamista. Jakson pituus vaihtelee urheilijan tason mukaan kahdesta viikosta jopa kuuteen viikkoon. Tärkein tavoite on fyysisesti ja psyykkisen väsymyksen poistaminen. Siirtymävaiheen aikana tulee olla aktiivinen, pelkkä passiivinen lepo ei välttämättä riitä palautumiseen. Suositeltavaa on myös antaa paljon käytetyille nivelille ja jänteille lepoa. Esimerkiksi työntökädelle ei tehdä siirtymäkaudella työntöjä (penkki, lajisuoritukset, pystypunnerrus ym.). Selän kompressio- ja leikkausvoimia selälle on lisäksi hyvä välttää (kyykky, maastaveto ja vastaavat liikkeet).
Viimeinen ominaisuuksien
nosto: herkistely
Herkistely on kilpailukaudella erittäin tärkeää. Tarkoituksena on poistaa harjoittelun aiheuttama väsymys mutta ylläpitää ja jopa nostaa esiin tiettyjä ominaisuuksia. Taulukosta 5 voi nähdä, että karkeasti harjoittelun volyymia kannattaa laskea ja intensiteettiä ei. Herkistelyn pituus on hyvä olla 8 - 14 päivän välissä ja harjoitustiheyttä ei kannata laskea. Sopiva voluumin lasku näyttäisi olevan noin puolet normaalista. Tämä riittää säilyttämään saavutetut ominaisuudet, mutta poistamaan väsymyksen.
TAULUKKO 5: Herkistelyn vaikuttaa
suorituskykyyn merkitsevästi meta-analyysin mukaan. N=koehenkilöiden määrä ja
P=tilastollinen merkitsevyys (tilastollisesti merkitsevimmät tulokset
lihavoitu). Tehokkain tapa herkistellä on laskea harjoitusvolyymia 41-60 % mutta
samalla pitää intensiteetti ylhäällä. Kestoltaan 8-14 vuorokauden
progressiivinen herkistely näyttäisi olevan pituudeltaan tehokkain. Mukailtu
(Bosquet et al., 2007).
Lähteet:
Auvinen, M. 2011. SUL koulutusohjeet 2011-2013. s. 58-70. Koonnut Rajala, T. 2011. Viitattu 3.11.2016.
Bompa, T. & Haff, G. 2009. Periodization - Theory and methodology of training. 5.
painos. Human Kinetics.
Bosquet, L.,
Montpetit, J., Arvisais, D. & Mujika I. 2007. Effects of tapering on
performance: A Meta-analysis. Medicine & Science in Sports
& Exercise, 39 (8), 1358-1365.
Häkkinen
& Ahtiainen. 2016. Maksimivoimaharjoittelu. Teoksessa Mero, A., Nummela,
A., Kalaja, S. & Häkkinen, K. (toim) Huippu-urheiluvalmennus. 1.painos. VK-Kustannus Oy, 250-265.
Krieger, J. 2010.
Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy: A
Meta-Analysis. Journal of strength and conditioning research, 24 (4),
1150-1159.
Mero, A., Nummela,
A., Keskinen, K. & Häkkinen, K. 2007. Urheiluvalmennus.
2.painos. VK-kustannus.
Rooney, K., Herbert, R. & Balnave, R. 1994. Fatigue contributes to the strength training stimulus.
Medicine
and science in sports and exercise, 26 (9), 1160-1164.
Räntilä, A. 2015. Yleisurheilun heittolajien fyysiset
suorituskykyvaatimukset käytännön kenttätesteissä ja niiden vaikutus
kilpailutuloksiin. Haaga-Helia, opinnäytetyö.
Schoenfeld, B.
2013. Potential mechanisms for a role of metabolic stress in hypertrophic
adaptations to resistance training. Sports medicine, 43 (3), 179-194.
Schiaffino, S., Dyar, K., Ciciliot, S., Blaauw, B. &
Sandri, M. 2013. Mechanisms regulating skeletal muscle growth and atrophy. Federation of European Biochemical
Societies, 280,
4294–4314.
Schott, J.,
McCully, K. & Rutherford, O. 1995. The role of metabolites in strength
training. II. Short versus long isometric contractions. European journal of
applied physiology and occupationally physiology, 71 (4), 337-341.
Yrjölä, M. 2000. Suomalainen Kuulantyöntö. Lahtiprint
- Lahti.
Zatsiorsky, V. & Kraemer,
W. 2006. Science and Practice of Strength Training. 2. painos. Human Kinetics.
Laihduttaminen
on suhteellisen helppoa. Usein ongelmaksi kuitenkin muodostuu se, että miten
paino pidetään poissa. Laihduttajista vain 10-20% pystyvät pitämään painon
poissa laihdutuksen jälkeen (Wing & Phelan, 2005)! Miksi uudessa painossa
pysyminen on niin hankalaa?
Keho
käsittelee laihdutuskuureja kuten nälkiintymistä. Kun laihdutat, niin myös aineenvaihdunta
tippuu alas. Aineenvaihdunnan tippumisen tarkoituksena on vähentää
energiankulutusta, jotta nälkiintymisestä selviydytään. Evolutionaarisesta
näkökulmasta tämä on täysin järkevää, koska ruokaa ei ole ollut aina saatavilla.
Aineenvaihduntaa tulee voida säädellä, jotta selvitään erilaisista tilanteista.
Aineenvaihdunnan vähentämisen tarkoitus on vähentää kalorikulutusta, jotta
säästetään kallisarvoista energiaa. Ideana on suojella kehoa ja nykyistä
rasvaprosenttia. Keho pyrkii aina tasapainoon. Kun tasapaino heilahtelee, se
pyritään korjaamaan mahdollisimman nopeasti. Tämä ihmeellinen kykymme on
muuttunut nykymaailmassa ongelmaksi. Kaiken tämän lisäksi painon laskiessa
nälkä lisääntyy, joka johtaa ruoan etsimiseen ja siihen, että painoa saataisiin
taas takaisin ylöspäin. Kaikki tämä hankaloittaa laihduttamista.
Paras ratkaisu aineenvaihdunnan ylläpitämiseen on rauhallinen laihdutustahti ja pienet tauot dieeteistä. Taukojen aikana syödään kaloreita vain energiankulutuksen verran. Toimivin strategia on pitkäaikainen laihdutus ja elämäntapamuutos. Kesän alla moni ihminen haluaa yhtäkkiä laihtua 20 kiloa. Myös painoluokkaurheilijat sortuvat samaan laihduttaessaan äärimmäisen nopeasti kisojen alla jopa kymmenenkin kiloa. Toisaalta voisi miettiä, kuinka kauan lihomiseen meni? Jos aikaa meni vaikka seitsemän vuotta, niin voiko kuvitella painon lähtevän parissa kuukaudessa pois? No ei voi. Karkeasti painon pudottamiseen voisi varata kolmasosan siitä ajasta mikä meni painon kertymiseen. Seuraavaksi katsotaan, miksi laihdutuskuurit ovat niin vaikeita ja mitä fysiologia sanoo laihduttamisen mekanismeista?
Miksi
olemme lihoneet?
Maailma on muuttunut, ja ne asiat, jotka nyt hankaloittavat laihduttamista, olivat ennen pelastuksemme. Ruokaa oli tarjolla vain harvoin, ne yksilöt, jotka onnistuivat säilömään sitä pahan päivän varalle, olivat vahvoilla. Ihmisen kyky varastoida rasvaa on auttanut esi-isiämme selviytymään sellaisina aikoina, kun ruokaa ei ollut joka päivä saatavilla. Lisäksi ihmisen kyky säädellä aineenvaihduntaa on auttanut meitä selviytymään ruoattomista vaiheista. Kun ruokaa ei ole saatavilla, pystymme kuluttamaan vähemmän kaloreita eli säätämään aineenvaihduntaamme. Nykyään ruokaa on koko ajan tarjolla, ja tämä etu on muuttunut ongelmaksi. Ohessa kolme kuvaa, joilla voidaan hiukan selittää sitä, miksi ihmiset lihovat enemmän ja miksi laihduttaminen on tärkeä aihe yhteiskunnallisesti.
Lihavuustutkija Stephan J. Guyenet on tehnyt upean kuvaajan lihavuudesta ja energian lisääntymisestä. Lihavuus kuvattuna punaisella ja harmaalla ovat nousseet viime vuosikymmeninä samaan tahtiin kuin ruoan kulutus (vihreällä). 1970-luvulta saakka lihavuuden esiintyvyys Yhdysvalloissa on jopa tuplaantunut! Noin 35% amerikkalaisaikuisista on lihavia. Samaan aikaan amerikkalaiset syövät noin 400 kaloria enemmän päivässä kuin ennen. Kasvu on viidenneksen luokkaa. (Guyenet, 2014. Data: CDCNHANES – kyselystä.)
Myös amerikkalaisten syömiskulttuuri on muuttunut. Amerikkalaiset syövät yhä enemmän muualla kuin kotona, ja tämä saattaa selittää lisääntyneitä kalorimääriä päivässä. (Pereira et al., 2005.) Yhdysvaltalaisten kalorien kulutus yhden päivän aikana on myös vähentynyt dramaattisesti. Yksi tärkeä tekijä on työn ja yhteiskuntamme suuri muutos. Liikumme työssä koko ajan yhä vähemmän ja kulutamme vähemmän kaloreita. Työ on vähemmän fyysistä kuin ennen, mutta ruokailutapamme eivät ole muuttuneet työn muutoksen myötä, päinvastoin. (Church et al., 2011.)
Nälän
ja aineenvaihdunnan lyhyt oppimäärä
Lipidit
eli rasva-aineet ovat kemiallisia yhdisteitä, ja ne voidaan jaotella
varastorasvaan sekä muutamaan vähemmän tärkeään rasva-aineeseen. Varastorasvoja
ovat triglyseridit, fosfolipidit ja kolesterolit. Keskitymme erityisesti
triglyserideihin, koska juuri niihin varastoinut rasva on yleensä se, jota
halutaan vähentää. Triglyseridi muodostuu glyserolista ja siihen liittyneestä
kolmesta rasvahaposta, siitä nimi tri – kolme ja glyseridi – glyserolin esteri.
Suuri määrä rasvaa on varastoitu rasvakudokseen
ja maksaan. Rasvakudoksen tärkein tehtävä on varastoida triglyseridejä, kunnes
niitä tarvitaan energiantuottoon muualla elimistössä. Se toimii myös
lämmöneristäjänä kehossa. Rasvakudoksen rasvasolut varastoivat triglyseridejä
jopa 85-90 % koko solun tilavuudesta. Rasvasolut säilövät triglyseridit yleensä
nesteen muodossa, sillä vain nestemäinen rasva voidaan hydrolysoida ja
kuljettaa soluista toiseen
Jos varastorasva halutaan ottaa käyttöön energiaksi, ensimmäinen vaihe triglyseridien käytössä on niiden hydrolyysi (kemiallinen reaktio, jossa yhdiste hajoaa) rasvahapoiksi ja glyseroliksi. Tämän jälkeen rasvahapot ja glyseroli kuljetetaan veressä aktiivisiin kudoksiin, jossa ne hapetetaan energian tuottamiseksi. Glyseroli voidaan melkein suoraan käyttää energiaksi glykolyyttisessä reitissä. Rasvahappojen hajoaminen ja hapetus vaatii puolestaan hiukan pidemmän reitin. Rasvahappojen käytön ensimmäinen vaihe on siirtyminen mitokondrioihin, jossa karnitiini toimii kantaja-aineena. Mitokondrioissa rasvahapot irtoavat karnitiinista ja sen jälkeen hapettuvat ja hajoavat. Rasvahappomolekyyli hajotetaan mitokondriossa asetyylikoentsyymi-A:n asteittaisen vapauttamisen avulla. Tätä tapahtumasarjaa kutsutaan β-oksidaatioksi. Laihduttaminen johtaa suuriin muutoksiin mitokondrion toiminnoissa. Tämän takia on tärkeä ymmärtää mitokondrion rooli rasva-aineenvaihdunnassa.
Kun keho muodostaa varastorasvaa ylimääräisestä energiasta, kyseessä on harvinaisen tehokas prosessi. Triglyseridien synteesin aikana vain noin 15 % glukoosin alkuperäisestä energiasta häviää lämpönä, loput 85 % siirretään varastoituihin triglyserideihin. Rasvan synteesi hiilihydraateista on erityisen tärkeää, koska kehon solujen kyky varastoida hiilihydraatteja glykogeenin muodossa on yleensä vähäinen. Kaiken kaikkiaan vain muutama sata grammaa glykogeenia voidaan varastoida maksassa, luustolihaksissa ja kaikissa muissa kehon kudoksissa yhteensä. Sen sijaan rasvakudokseen voidaan varastoida monta kiloa rasva. Rasvasynteesi tarjoaa keinon, jolla ylimääräiset hiilihydraatit ja proteiinit voidaan varastoida myöhempää käyttöä varten. Lisäksi jokainen gramma rasvaa sisältää lähes 2,5 kertaa enemmän energiaa kuin gramma glykogeenia. Tietyllä painon nousulla voidaan varastoida useita kertoja enemmän energiaa rasvan muodossa kuin hiilihydraatteina.
Rasvasolulla on muitakin tehtäviä kuin pelkkä rasvan varastointi. Nykytietämyksen valossa rasvasolu ei ole enää vain yksinkertainen varastosolu, vaan se toimii monimutkaisen elimen tavoin. Ne kommunikoivat hormonien avulla, lähettävät signaaleja aivoille ja yhdistävät monia muita signaaleja. Näin rasvasolu on aktiivinen toimija, ei pelkkä varastosolu.
Rasvasolu voidaan nähdä ikään kuin termostaattina. Se säätelee rasvan määrä kehossa. Jos rasvaprosentti nousee, niin leptiinitasot nousevat myös. Leptiini on rasvasolujen tuottama hormoni, jonka tarkoitus on säädellä ihmiskehon varastorasvan määrää. Leptiinin rooli laihdutuksessa on suuri. Kun rasvasolut ovat ”täysiä”, niin leptiiniä tuotetaan paljon. Tämä viestii aivoille, että emme tarvitse enempää ravintoa vaan olemme kylläisiä. Kun taas laihdumme, ja rasvasolut varastoivat vähemmän energiaa kuin ennen, ne tuottavat vähemmän leptiiniä. Leptiini kulkeutuu aivoihin hypotalamukseen, jossa mm. kontrolloidaan nälkää ja aineenvaihduntaa. Hypotalamuksen leptiinireseptorit auttavat rasvasoluja kommunikoimaan suoraan aivoille. Rasvasolut huutavat siis pienentyessään, että tarvitaan lisää ruokaa! Korkeat leptiinitasot aktivoivat sympaattista hermostoa hypotalamuksen kautta, mikä johtaa aineenvaihdunnan kiihtymiseen. Muun muassa ruskean rasvan aineenvaihdunta kiihtyy, ja samalla lämmöntuotto suoraan lihaksissa lisääntyy.
Nälän
säätelyssä hypotalamuksen rooli on äärimmäisen tärkeä. Hypotalamuksessa
vaikuttavat hermovälittäjäaineet ja hormonit voidaan jakaa nälän tunnetta
aiheuttaviin ja kylläisyyden tunnetta aiheuttaviin. Hypotalamukseen saapuu
signaaleja ympäri kehoa. Nälän säätelyyn vaikuttavat muun muassa seuraavat
viestit:
Aivojen kuorikerroksesta tulevat haju-, näkö- ja
makuaistimukset.
Jos taas rasvaprosentti laskee, niin leptiinitasotkin laskevat, ja tämä aiheuttaa korostuneen nälän tunteen. Lisäksi aineenvaihdunta laskee, ja nämä muutokset aiheuttavat rasvaprosentin nousun takaisin alkutilanteeseen. Rasvasolu on toiminut termostaatin tavoin. (Pandit et al., 2017.)
Lihoessa taas aineenvaihduntaa pyritään kiihdyttämään, jotta rasvaprosentti saataisiin takaisin alkutilanteeseen. Kaikki kunnon bulkilla (massakuurilla) olleet tietävät sen tunteen, kun ruoka ei enää maistu, ja pelkkä ajatus syömisestä oksettaa. Tämä on seurasta leptiinitasojen noususta. Rasvasolu on erittänyt lisää leptiiniä, jotta hypotalamus osaisi säädellä nälän tunnetta, ja rasvasoluun ei tuotaisi enää yhtään lisää rasvaa.
Laihtumisen kannalta myös kilpirauhashormonit ovat tärkeitä. Kilpirauhanen tuottaa kahta aineenvaihdunnallisesti aktiivista hormonia, jotka ovat tyroksini ja trijodityroniini. Korkeat kilpirauhashormonitasot on yhdistetty korkeaan aineenvaihduntaan. Tämä johtuu siitä, että kilpirauhashormonit stimuloivat ATP:n kulutusta. Kun kulutus kasvaa, niin tuottoakin nostetaan, mikä johtaa suurempaan kalorien kulutukseen. (Harper & Seifert, 2008.) Lisäksi kilpirauhashormoni stimuloi ruskean rasvan aktiivisuutta. Ruskea rasva tuottaa lämpöä ja täten kuluttaa lisää kaloreita. Kuitenkin ruskean rasvan rooli kokonaisuudessa on pieni. (Nedergaard., 2007.) Laihduttaminen kuitenkin laskee kilpirauhashormonitasoja (Pardue et al., 2017; Agnihothri, R., 2014). Kilpirauhashormonien lasku on yksi iso osa laihduttamisen aikaansaamia muutoksia, ja tämäkin muutos lisää rasvan varastointipotentiaalia.
Kortisoli lisää proteiinien hajotusta, ja täten rasvaton lihasmassa pienenee. Rasvattoman lihasmassan pieneneminen on yhdistetty valtavaan nälän tunteeseen, kunnes lihasmassa on palautunut (Dulloo et al., 2018). Muita tärkeitä hormoneita, jotka vaikuttavat nälän tunteeseen ovat greliini ja insuliini. Greliini stimuli ruokahalua ja nopeuttaa mahalaukun tyhjenemistä. Sitä erittyy mahalaukun limakalvolta mahalaukun venyessä. Insuliini puolestaan säätelee sokeriaineenvaihduntaa.
Leptiini
välittää aivoille tietoa elimistön rasvavarastojen määrästä. Leptiini kulkeutuu
aivoihin verenkierron välityksellä. Se muuttaa kehon kylläisyysolotilaa ja
säätelee painoa pitkällä tähtäimellä. Matala leptiinipitoisuus tekee
painonhallinnan vaikeaksi, koska se lisää nälkää ja vähentää kylläisyyden
tunnetta. Laihduttaminen laskee leptiinitasoja. Välillisesti leptiini lisää kilpirauhashormonin,
lisääntymishormonien, nälkähormonien ja kortisolin eritystä (Kelesidis, 2010).
Miksi
ylipaino palaa laihdutuskuurien jälkeen?
Jojo-laihdutus tarkoittaa kehon painon
toistuvaa laskua ja nousua laihduttamisen takia. Termillä kuvataan tilannetta,
jossa laihduttaja onnistuu pudottamaan painoaan, muttei onnistu säilyttämään
pudonnutta painoa. Painon nousun jälkeen laihduttaja pyrkii jälleen pudottamaan
uudelleen painoa.
Ihmistutkimuksia
aiheesta on vähän johtuen siitä, että kontrolloidussa ympäristössä ihmisten
lihottaminen ja laihduttaminen vuoron perään on eettisesti arveluttavaa.
Rotille on tehty suhteellisen paljon jojo-laihdutustutkimuksia. Laihdutuksen
jälkeen painon takaisin saaminen on tapahtunut puolessa ajassa siitä, mikä
niillä meni tiputtaa se. Kun taas laihdutettiin, rotat tiputtivat painoa kaksi
kertaa hitaammin kuin aikaisemmin. Kun rotat saivat uudelleen syödä, niin niiden
paino nousi kolme kertaa nopeammin kuin aluksi. Jyrsijöistä tuli siis joka
syklin jälkeen tehokkaampia varastoimaan rasvaa! Aina kun rotat menivät tallaisen
laihdutussyklin läpi, niiden aineenvaihdunta hidastui. Rotat eivät olleet enää
yhtä tehokkaita laihtumaan, vaan niistä tuli yhä tehokkaampia varastoimaan rasvaa
takaisin kehoon. (Brownell et al., 1986.)
Laihtuessa rasvasolujen halkaisija pienenee, ja lihoessa se kasvaa. Kuitenkin joskus rasvasolun koon kasvaessa tietyn halkaisijan yli (joissain tutkimuksissa on puhuttu 100 mikrometristä) voi syntyä uusia rasvasoluja. Tämä on ongelmallista, koska uudet rasvasolut mahdollistavat suuremman rasvavarastointipotentiaalin, ja tällöin mahdollisuus lihomiselle on suurempi kuin aikaisemmin. Rotilla tehdyissä tutkimuksissa rasvasolujen lukumäärä saattoi lisääntyä jopa 50%. Uusien rasvasolujen synnyttyä keskiarvo rasvasolun halkaisija voi yhä olla pienempi kuin ennen, joten keho voi luulla, että olet kalorivajauksella, vaikka olet samassa rasvaprosentissa kuin ennen! (Jackman et al., 2008.)
Tällainen tilanne kävi juuri Jackmanin ja hänen kollegoidensa (2008) tutkimuksessa. Rottien alkutilanteessa rasvasolun halkaisija oli noin 103 mikronia (1000 mikronia on 1 millimetri). Jyrsijöillä oli 37 miljoona rasvasolua, ja keskiarvopaino oli 709 g. Rotat laitettiin kalorivajauksella, ja paino tippui 583 grammaan, ja rasvasolujen koko pieneni 85 mikroniin. Laihdutus onnistui siis hyvin. Sen jälkeen rotat saivat syödä niin paljon kuin halusivat. Paino nousi 609 g, rasvasolun koko pysyi samana (80 mikronia), mutta rasvasolujen määrä pamahti 55 miljoonaan (+49%). Suurempi määrä rasvasoluja on paljon isompi potentiaali varastoida rasvaa. Kun rotat palasivat alkupainoonsa (700 g) oli rasvasolujen määrä sama 55 miljoonaa, mutta rasvasolun halkaisija vain 94 mikronia. Rasvasolujen halkaisija oli vieläkin alle alkutilanteen. Jokainen yksittäinen rasvasolu lähetti koostaan koko ajan signaaleja eteenpäin. Tämä johti siihen, että leptiiniä oli vähemmän, ja näläntunne pysyi, vaikka rotat olivat alkupainossaan! Arvaatteko mitä tapahtui? Rottien paino pysähtyi 740 grammaan, ja rasvasolun halkaisija palasi täsmälleen samaan 103 mikroniin, missä se oli alussakin. Mutta koska rasvasoluja oli 55 miljoonaa, niin painoa kertyi enemmän (+ 31 g) kuin alkutilanteessa. Rottien aivot luulivat, että ollaan samassa tilanteessa kuin alussa, vaikka painoa oli enemmän kuin aluksi (+ 4,4%).
Rasvasolujen määrä lisääntyy, ja rasvasolut palaavat alkutilanteessa olleeseen halkaisijaansa. Rasvasolut ovat siis samankokoisia kuin ennen, mutta niitä on enemmän. Teoreettinen malli, jonka on kehittänyt Peos et al., 2019. Kuvan lähde sama.
Kun
rasvaprosentti nousee, leptiinin erityskin lisääntyy. Tämä johtaa kohonneeseen
aineenvaihduntaan ja kylläisyyden tunteeseen. Nämä lopulta johtavat syömisen
vähentymiseen ja rasvaprosentin laskuun. Keho puolestaan puolustaa painoaan ja
haluaa palata siihen takaisin edellä mainittujen reaktioiden kautta. Jos taas
kehon rasvaprosentti laskee, leptiinin erityskin vähenee. Tämä johtaa
pienentyneeseen aineenvaihduntaan, jotta kulutus vähenee, ja kohonneeseen nälän
tunteeseen, jotta ravintoa syötäisiin lisää. Pitkällä aikavälillä tämä johtaa
rasvaprosentin nousuun. Leptiini toimii tässä tärkeimpänä välittäjähormonina.
Yhdysvalloissa Suurin Pudottaja –ohjelmaan osallistuneista kilpailijoista on tehty mielenkiintoinen tutkimus. Ohjelman ideana on pudottaa painoa mahdollisimman nopeasti ja äärimmäisillä keinoilla. Kilpailijat harjoittelevat ohjelman aikana yhtenä päivänä jopa 4-6 kertaa ja syövät erittäin rajoitetusti. Tulokset ovat olleet huikeita, yksi kilpailun osallistujista, Danny Cahill, tiputti painoa 195 kilosta aina 86 kiloon. Tämä on kaikkien aikojen ennätys ohjelman aikana, yhteensä painoa tippui huimat 108 kg. Seuraavan kuuden vuoden aikana hän lihoi 45 kg takaisin. Miksi näin tapahtuu?
Tutkijat (Fothergill et al., 2016) seurasivat 14 (6 miestä ja 8 naista) ohjelmaan osallistunutta henkilöä kuusi vuotta ohjelmaan osallistumisen jälkeen. Tulokset näkyvät koottuna Taulukossa 1 ja ovat aika uskomattomia. Vain yksi henkilö ei ollut lihonut yhtään takaisin. Keskiarvollisesti lähes kaikki palasivat takaisin alkupainoonsa, ja neljä painoi enemmän kuin ennen ohjelmaan osallistumista. Kaikkein mielenkiintoisin löytö oli kuitenkin se, että heidän aineenvaihduntansa oli hidastunut dramaattisesti painonpudotuksen myötä. Keskiarvollisesti kalorinkulutus tippui ohjelman aikana 2607 kalorista 2000 kaloriin! Kuuden vuoden jälkeen tämä oli tippunut keskiarvollisesti jo 1903 kaloriin päivässä! Aineenvaihdunta tulee siis laskemaan laihdutuksen myötä. Tämä tulee huomioida laihdutussuunnitelmassa.
Jotkut Suurimpaan Pudottajista osallistuneista ihmisistä lihoivat takaisin lähtöpainoonsa ja jopa sen ylikin.
Miten
ehkäistä negatiivisia muutoksia ja onnistua laihdutuksessa?
Onko mitään järkeä edes yrittää laihduttaa, jos painon kuitenkin saa heti takaisin? On, mutta tarvitaan pitkäaikainen suunnitelma. Nopeat ratkaisut laihduttamisessa eivät johda hyviin tuloksiin. Pitkällä aikavälillä ja maltillisella painonpudotuksella pystytään välttämään takaisin lihominen. Pitkällä aikavälillä kehon signaalit eivät ole niin voimakkaita, ja niitä pystytään kontrolloimaan paremmin, kun laihdutus ei ole äärimmäistä. On myös jotain aavistelua siitä, että rasvasolu tottuisi uuteen kokoonsa muutaman vuoden aikana eikä enää lähettäisi signaaleja ”väärästä” koostaan. Tämä adaptoituminen tapahtuu pitkällä aikavälillä, ja myös sen takia on tärkeä tehdä laihdutussuunnitelma pitkällä aikavälillä.
Laihtuessa
aineenvaihduntasi tulee laskemaan. Aineenvaihdunnallisia muutoksia tulee
tapahtumaan laihdutuksen aikana, eikä niitä voi estää. Pienempi versio sinusta
tarvitsee loppuelämän vähemmän kaloreita kuin ennen. Elämäntapamuutos kestää
loppuelämän, ei laihdutuskuurin verran. Tämän huomioiminen
laihdutussuunnitelmassa on äärimmäisen tärkeätä!
Aineenvaihdunnallisia muutoksia voi ennaltaehkäistä seuraavilla tavoilla. Karkean arvion mukaan proteiinia tulisi saada 2 grammaa painokiloa kohden päivässä. Urheilijoilla proteiinin saanti voi olla jopa 2.3-3.1 grammaa painokiloa kohden riippuen lajista/tavoitteista. Esimerkiksi satakiloinen henkilö voisi syödä jopa 200g-230g proteiinia päivässä ehkäistäkseen lihasten kataboliaa ja nälän tunnetta. Proteiini vähentää mm. greliinin eritystä, joka lisää nälän tunnetta. Proteinilla on huomattavasti suurempi terminen vaikutus kuin rasvalla tai hiilihydraatilla, joten sen voidaan ajatella lisäävan ainenvaihduntaa.
Lisäksi aineenvaihdunnallisia muutoksia voi ehkäistä maltillisella painonpudotuksella. Painonpudotus tulisi olla maksimissaan 1 % kehonpainosta viikossa, mieluiten 0,25-0,5 % viikossa. Kuulostaako tututulta? Ei minustakaan, harva laihduttaja tekee näin maltillisia tavoitteita. Pienet rasvaprosentin muutokset ovat todella paljon helpompia ylläpitää! Esimerkiksi ensiksi rasvaprosentin pudotus 18 prosentista 16 prosenttiin, sitten ylläpitokaloreita parin kuukauden ajan ja vasta sitten pudotus 16 prosentista 14 prosenttiin. Jos olet painoluokkaurheilija, pysy lähellä omaa painoluokkaasi! Painon kanssa jojoilu johtaa suurempaan painoon keski-iässä. Tämä on seurasta juuri rasvasolujen hyperplasiasta. (Saarni et al., 2006.)
Lähes
kaikki dieetit voivat toimia, kunhan ne luovat kalorivajauksen. Tärkeimpänä on
valita sellainen käytösmalli, joka on helppo ja toimiva itselle. Tauot dietistä 1-2 viikkoa kerrallaan voivat ehkäistä
aineenvaihdunnallisia muutoksia mitokondrioiden toiminnoissa, hormonien
erityksessä ja perusaineenvaihdunnassa. Tällaisilla tauoilla on tarkoitus syödä
kaloreita ylläpitomäärän verran, ei ahmia niin paljon kuin haluaa.
Liikunnalla
on suuri vaikutus. Toinen artikkeli, joka on julkaistu liittyen samaan Suurin pudottaja
-tutkimukseen, kertoo, että ne, jotka olivat nostaneet päivittäistä
aktiviteettia 160 %, olivat onnistuneet pitämään painonsa kurissa paremmin. Ne,
jotka olivat lihoneet lähemmäksi alkutilannetta, olivat lisänneet päivittäistä aktiviteettia
vain 34 % entiseen elämäänsä verrattuna (Kerns et al., 2017). Fyysisellä
aktiivisuudella on siis tärkeä rooli, jotta paino saadaan pidettyä poissa. Lisäksi
fyysinen harjoittelu lisää rasvojen hapettamista rasvasoluista ja tuottaa
lukemattomia muita positiivisia reaktioita.
Unella
ja stressillä on myös valtaisa vaikutus painoon. Unen puute aiheuttaa
metabolisia ja hormonaalisia muutoksia muun muassa huonontaen
glukoositoleranssia ja insuliinisensitiivisyyttä, laskien leptiinitasoja, lisäten
nälkää ja ruokahalua ja lisäten kortisolin sekä reliinin eritystä (Leptroult
& Van Cauter, 2010). Väsyneenä haluaa myös syödä kaloritiheämpiä ruokia, ja
näin kalorien saanti voi kasvaa huomaamattakin. Ne yksilöt, jotka nukkuivat 5,5
tuntia yössä, menettivät enemmän lihasta ja vähemmän rasvaa kuin ne, jotka
nukkuivat 8,5 tuntia yössä (Nedeltcheva et al., 2010). Unta voi parantaa
vähentämällä sinistä valoa ennen nukkumaan menemistä. Lisäksi kofeiinin ja
alkoholin vähentäminen parantaa unen laatua. Hätäkeinona hiilihydraattien
painottaminen iltaan voi auttaa nukahtamista ja rauhoittumista.
Lisäksi stressi voi johtaa unen lyhenemiseen, kun taas unen puute voi johtaa stressiin. Nämä taas voivat vaikuttaa aineenvaihduntaan kortisolin kautta. (Han et al., 2012.) Tärkeä on siis luoda joustava laihdutussuunnitelma, joka ei stressaa laihduttajaa. Laihdutusstressi itsessään saattaa siis estää laihdutussuunnitelmasi onnistumisen!
Periytyykö
lihavuus? Syömishäiriöt ja biologiset signaalit?
Lapsuusajan lihavuus näyttäisi todellakin kantavan läpi elämän, ja se näyttäisi johtavan lihavuuteen myös aikuisiällä. Syynä tähän on etenkin lapsuusajan lihavuuden aikaansaama lisääntynyt rasvasolujen määrä.
Voidaan
myös sanoa, että lihavuus periytyy. On kuitenkin vaikea sanoa, kuinka suuri
osuus johtuu geeneistä. Tällä hetkellä tutkimus näyttää, että noin 20-25 % lihavuudesta
johtuisi geneettisistä syistä. Lihavuutta aiheuttavat geenit vaikuttavat
syömiskäyttäytymiseen vaikuttavilla aivojen alueilla sekä aineenvaihduntaan ja
rasvan varastointiin vaikuttavissa paikoissa. On pystytty identifioimaan kolme erilaista
yksittäistä geeniä, joiden eri muodot vaikuttavat lihavuuden syntyyn. Yksi
vaikuttaa MCR-4 reseptoreihin, toinen leptiinigeeniin ja kolmas
leptiinireseptoreihin. Nämä geenimuodot ovat kuitenkin erittäin harvinaisia
eivätkä ne selitä lihavuutta väestötasolla. (Guyton & Hall, 2011.)
Kuten olemme aikaisemmin huomanneet, leptiini on yksi tärkeimmistä tekijöistä laihdutuksessa. Mitä jos leptiiniä ruiskuttaisi suoraan suoneen? Vältettäisiinkö näin suurin osa ikävistä haittavaikutuksista? Alustavasti näyttäisi siltä. Leptiinilisäys on tutkimuksissa havaittu toimivaksi ratkaisuksi (Rosenbaum & Leibel, 2013). Ikävä kyllä tämä oikotie ei onnistu käytännössä. Leptiini on todella kallista, sivuvaikutuksia ei tunneta, ja käytännössä sitä ei saa mistään. Tulevaisuudessa leptiiniruiskutus voi olla yksi vaihtoehto laihduttaessa. Toisaalta leptiinin liikaeritys on myös yhdistetty syöpään, diabeteksen ja sydän- ja verisuonitauteihin. Voi siis mennä hetki ennen kuin leptiinilääke nähdään apteekeissa.
Pitää myös mainita, että syömishäiriöt muuttavat ihmisen tuntemia nälkään liittyviä signaaleja. Syömishäiriöinen kykenee sulkemaan ja sivuuttamaan kaikki aivoihin tulevat signaalit ruoan suhteen tai muuttamaan niitä voimakkaasti. Käsittelemämme teoria koskeekin vain niitä, joilla on jokseenkin normaali suhtautumistapa ruokaan.
Yhteenveto
Rasvasolu on muutakin kuin varastosolu. Se toimii aktiivisena toimijana välittäen tietoa aivoille. Laihduttaminen johtaa rasvasolun pienentymiseen. Tällöin rasvasolu lähettää jatkuvasti tietoa aivoille, että ruokaa pitäisi saada lisää. Tämän näläntunteen kontrollointi voi olla vaikeaa. Tahdonvoimalla ei ole mitään tekemistä laihdutuksen kanssa, koska biologiset signaalit lisäävät koko ajan syömishalua, ja niitä on lopulta lähes mahdoton koko ajan vältellä. Maltillinen laihduttaminen ehkäisee näitä signaaleja ja ennen kaikkea takaisin lihomista.
Laihtuessasi
tapahtuu monia aineenvaihdunnallisia muutoksia. Huomioi nämä muutokset
laihdutussuunnitelmassa. Tauot laihdutuskuurista, maltillinen suunnitelma ja
riittävä proteiinin saanti voivat auttaa ehkäisemään aineenvaihdunnallisia
muutoksia.
Lähteet:
Agnihothri, R., Courville, A.,
Linderman, J., Smith, S., Brychta, R., Remaley, A., Chen, K., Simchowitz, L.
& Celi, F. (2013) Moderate Weight Loss Is Sufficient to Affect Thyroid
Hormone Homeostasis and Inhibit Its Peripheral Conversion. Thyroid: 24(1):
19-26. 10.1089/thy.2013.0055.
Brownell, D., Greenwood, M.R.
Stellar, E. & Shrager, E.E. (1986). The effects of repeated weight loss and
regain in rats. Physiology & behavior. 38. 459-64.
10.1016/0031-9384(86)90411-7.
Church, T., Thomas, D.,
Tudor-Locke, C., Katzmarzyk, P., Earnest, C., Rodarte, R., Martin, C., Blair,
S. & Bouchard, C. (2011) Trends over 5 Decades in U.S. Occupation-Related
Physical Activity and Their Associations with Obesity. Plos one, https://doi.org/10.1371/journal.pone.0019657
Dulloo,
A., Miles-Chan, J. & Schutz, Y. (2018) Collateral
fattening in body composition autoregulation: its determinants and significance
for obesity predisposition. Eur J Clin Nutr;
72(5):657–664. 10.1038/s41430-018-0138-6
Fothergill, E., Guo, J., Howard, L., Kerns, J., Knuth, N., Brychta, R., Chen, K., Skarulis, M., Walter, M., Walter, P. & Hall, K. (2016) Persistent metabolic adaptation 6 years after “The Biggest Loser,” competition. Obesity (Silver Spring) 2016; 24: 1612- 1619.
Guyton. A. & Hall J.
(2011) Textbook of Medical Physiology. 12th edition. Philadelphia, USA: W.B.
Saunders Company.
Han,
K., Kim, L. & Shim, I. (2012) Stress and sleep
disorder. Exp Neurobiol, 21:141–50. 10.5607/en.2012.21.4.141
Harper, M. & Seifert, E.
(2008) Thyroid hormone effects on mitochondrial energetics. Thyroid; 18(2):
145-156.
Jackman, M. R., Steig, A.,
Higgins, J. A., Johnson, G. C., Fleming-Elder, B. K., Bessesen, D. H., &
MacLean, P. S. (2008). Weight regain after sustained weight reduction is
accompanied by suppressed oxidation of dietary fat and adipocyte
hyperplasia. American Journal of
Physiology-Regulatory, Integrative and Comparative Physiology, 294(4),
R1117-R1129.
Kelesidis, T. (2010) Narrative
Review: The Role of Leptin in Human Physiology: Emerging Clinical Applications.
Annals of Internal Medicine. 152. 93. 10.7326/0003-4819-152-2-201001190-00008.
Kerns, J., Guo, J., Fothergill, E., Howard, L., Knuth, N., Brychta, R., Chen,
K., Skarulis, M., Walter, P. & Hall, K. (2017) Increased
physical activity associated with less weight regain six years after “The
Biggest Loser,” competition. Obesity
(Silver Spring) 2017; 25: 1838- 1843.
Leproult, R. & Van Cauter,
E. (2010) Role of sleep and sleep loss in hormonal release and metabolism.
Pediatric Neuroendocrinology, 17: 11-21.
Nath. S. (2016). The
thermodynamic efficiency of ATP synthesis in oxidative phosphorylation.
Biophysical Chemistry, 219: 69-74.
Nedeltcheva, A., Kilkus, J.,
Imperial, J., Schoeller, D. & Penev, P. (2010) Insufficient Sleep
Undermines Dietary Efforts to Reduce Adiposity. Annals of internal medicine.
153. 435-41. 10.1059/0003-4819-153-7-201010050-00006.
Nedergaard, J., Bengtsson, T. &
Cannon, B. (2007) Unexpected evidence for active brown adipose tissue in adult
humans. American Journal of Physiology-Endocrinology and Metabolism: 293(2):
444-52.
Pandit, R., Beerens, S. &
Adan, R. (2017) The role of leptin in energy expenditure: The hypothalamic
perspective. American journal of physiology. Regulatory, integrative and
comparative physiology. 312. ajpregu.00045.2016. 10.1152/ajpregu.00045.2016.
Pardue, A., Trexler, E. &
Sprod, L. (2017) Case Study: Unfavorable But Transient Physiological Changes
During Contest Preparation in a Drug-Free Male Bodybuilder. International
Journal of Sport Nutrition and Exercise Metabolism. 27. 1-24.
10.1123/ijsnem.2017-0064.
Pereire, M., Kartashov, A.,
Ebbeling, C., Van Horn, L., Slattery, M., Jacobs. & Ludwig, D. (2005)
Fast-food habits, weight gain, and insulin resistance (the CARDIA study):
15-year prospective analysis. Lancet, 365: 36-42.
Peos, J., Norton, L., Helms,
E., Galpin, A. & Fournier, P. (2019) Intermittent Dieting: Theoretical
Considerations for the Athlete. Sports, 16;7(1). pii: E22. doi:
10.3390/sports7010022.
Rosenbaum, M. & Leibel, R.
(2010) Adaptive thermogenesis in humans. International journal of obesity: 34
Suppl 1. S47-55. 10.1038/ijo.2010.184.
Saarni, S., Rissanen, A.,
Sarna, S., Koskenvuo, M. & Kaprio, J. (2006) Weight cycling of athletes and
subsequent weight gain in middleage. Internatiol journal of obesity, 30(11):1639-44
Wing, R. & Phelan, S.
(2005) Long-term weight loss maintenance. American journal of clinical
nutrition, 82: 222-225. doi:
10.1093/ajcn/82.1.222S.
Olen pääkaupunkiseudulla vaikuttava voima-, fysiikka- ja yleisurheiluvalmentaja. Valmentajana olen toiminut kohta 10 vuoden ajan. Olen erityisen kiinnostunut urheilijoiden suorituskyvystä ja harjoittelun yksilöllisistä eroista urheilijoilla sekä periodisaatiosta. Saan urheilijat suoriutumaan lajissaan paremmin.
Tämä sivusto on luotu aputyökaluksi valmentajille, jotka haluavat perustaa valmentamisensa tieteelliseen näyttöön. Alalla on vielä paljon käytössä vanhentuneita toimintatapoja, jotka heikentävät urheilijoiden suorituskykyä. Pyrin jakamaan tieteellistä tietoa ja auttamaan objektiivisesti kaikkia valmentajia.