Kuukautiskierron vaikutus urheilusuorituksiin on monisyinen, mutta tieteellinen näyttö suorituskyvyn vaihtelusta eri vaiheissa on ristiriitaista.
Harjoittelua ei kannata liikaa periodisoida pitkällä aikavälillä kuukautiskierron mukaan.
Hormonaalinen ehkäisy voi lievittää kierron oireita, mutta sen vaikutus suorituskykyyn on usein neutraali tai hieman negatiivinen.
Korkealaatuista tutkimusta tarvitaan lisää yksilöllisten erojen ja lyhytaikaisten hormonaalisten vaihteluiden vaikutusten ymmärtämiseksi.
Energiavaje ja kuukautiskierron häiriöt, kuten amenorrea, voivat merkittävästi haitata terveyttä ja suorituskykyä sekä selittää kuukautiskierron aikaista suorituskyvyn laskua.
Valmentajien ja urheilijoiden välinen avoin keskustelu sekä moniammatillinen tuki ovat avainasemassa kuukautiskierron vaikutusten hallinnassa.
Lukuaika: n. 7–8 minuuttia
Kuukautiskierron vaikutukset urheilusuorituksiin
Urheilevilla naisilla havaitaan usein kuukautishäiriöitä, joita ovat muun muassa ovulaation puuttuminen, luteaalivaiheen lyheneminen sekä kuukautisten harventuminen tai niiden poisjääminen (amenorrea). On arvioitu, että yli puolet aktiivisesti liikkuvista ja urheilijanaisista kärsivät jonkinlaisesta kuukautiskiertoon liittyvästä häiriöstä. Näiden taustalla voi olla elimistön kuukautiskiertoa säätelevien hormonipitoisuuksien aleneminen, joka voi johtua esimerkiksi kulutukseen nähden alhaisesta energiansaannista, liiallisesta harjoittelusta tai kokonaiskuormituksesta ja/tai fyysisestä tai psyykkisestä stressistä. On ratkaisevan tärkeää, ettei kuukautiskierron häiriöitä pidetä normaalina, vaan niitä tutkitaan ja hallitaan moniammatillisesti koko urheilijan uran ajan. Usein esimerkiksi kuukautisten pois jäämistä pidetään normaalina osana kovaa harjoittelua, mutta tämä ei ole totta. Avoin keskustelu kuukautisista auttaa valmentautumisessa huomioimaan paremmin kuukautiskierron mahdolliset yksilölliset vaikutukset urheilijan suorituskykyyn.
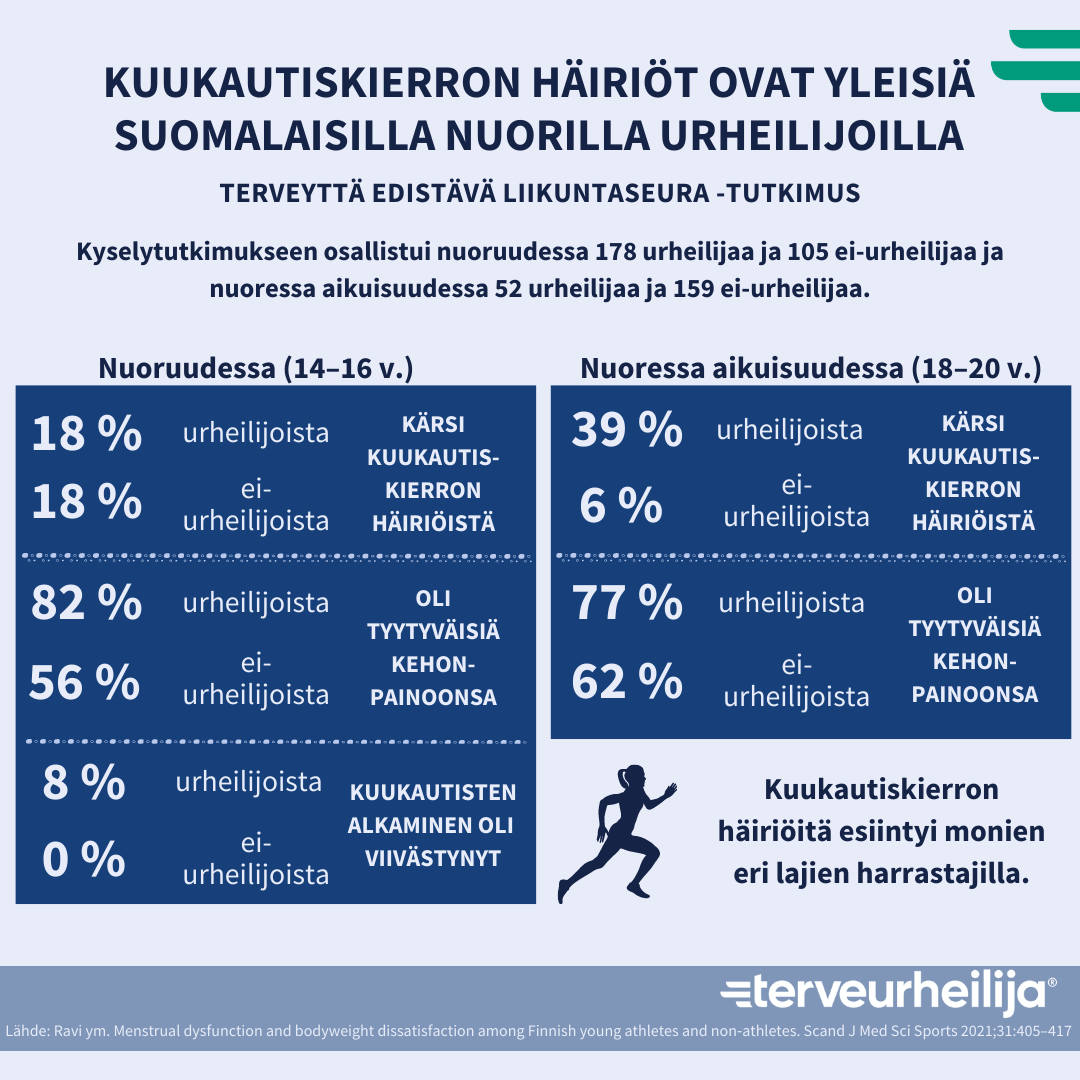
Kuva 1: Kuukautiskierron häiriöt ovat hyvin yleisiä suomalaiilla nuorilla urheilijoilla. Lähde: Ravi, S., Waller, B., Valtonen, M., Villberg, J., Vasankari, T., Parkkari, J., Heinonen, O. J., Alanko, L., Savonen, K., Vanhala, M., Selänne H., Kokko, S. & Kujala, U.M. Menstrual dysfunction and body weight dissatisfaction among Finnish young athletes and non-athletes. Scandinavian Journal of Medicine and Science in Sports 2021;31:405–417. Infograafi: terveurheilija.fi
Naisurheilijan hormonitoiminta
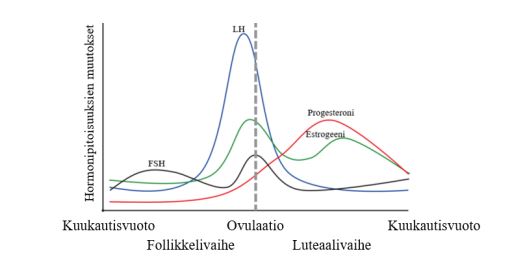
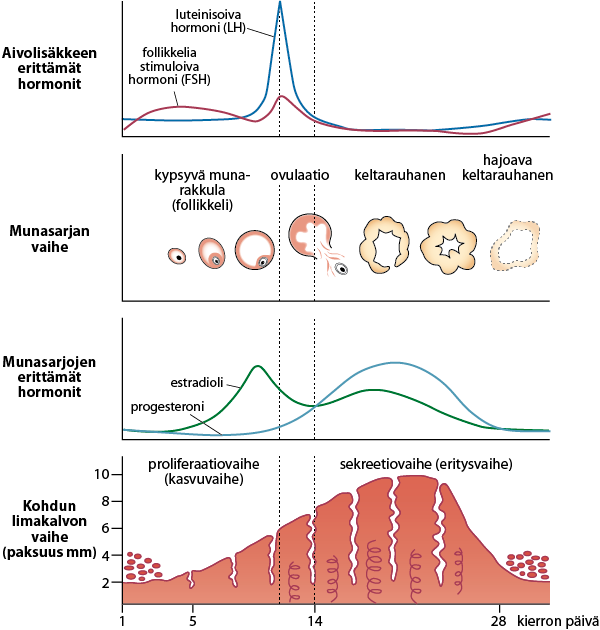
Kuukautiskierto jaetaan tavallisimmin kolmeen vaiheeseen hormonipitoisuuksissa tapahtuvien muutosten mukaan. Kaksi keskeistä kuukautiskiertoon liittyvää hormonia ovat estradioli (yksi estrogeenin esiintymismuodoista ja tässä tekstissä käytetään pitkälti estrogeeni-nimitiystä) ja progesteroni eli keltarauhashormoni. Estradiolin ja keltarauhashormonin pitoisuuksien vaihtelulla kuukautiskierron aikana on huomattavia vaikutuksia naisurheilijan fysiologiaan.
Androgeenihormoneihin lukeutuvan sukupuolihormonin testosteronin vaikutukset ovat erittäin hyvin tutkittuja, mutta estrogeenin ja progesteronin vaikutukset lihasmassan säätelyyn ovat heikommin tiedossa. Erilisissä tutkimuksissa on kuitenkin osoitettu estrogeenin vaikuttavan voimaharjoittelun adaptaatioihin, kuten proteiinin synteesiin ja myös degradaatioon, myosiinin toimintoihin sekäs satelliittisolujen aktiivisuuteen (18, 19). Näillä hormoneilla onkin suuri vaikutus voiman ja lihasmassan kehittymiseen pitkällä aikavälillä.
Lisäksi estrogeenin puute ja estrogeeni reseptorisignaloinnin vähentyminen vähentää satelliittisolujen määrää lihaksissa (19, 20), mikä saattaa vauhdittaa lihasmassan ja voiman vähentymistä esimerkiksi vaihdevuotisilla naisilla. Tämä tukee ajatusta siitä, että naisurheilijalla myös estrogeenillä on iso vaikutus voiman ja lihasmassan säätelyssä.
Kuukautiskierron suhteen estrogeenitasot ovat korkeimmillaan myöhäisessä follikulaarivaiheessa ennen ovulaatiota, kun taas progesteroni on korkeimmillaan ovulaation jälkeen puolessa välissä luteaalivaihetta. Tiedämme hormonien pienentymisen pitkällä aikavälillä olevan tuhoisaa lihas- ja voimaharjoittelun näkökulmasta, mutta kuukautiskierto vaikuttaa hormoneihin vain muutamien päivien ajan. Onko tällä lyhytaikaisella hormonivaihtelulla merkittävää vaikutusta akuuttiin suorituskykyyn tai pitkäaikaisiin hermolihasjärjestelmän adaptaatioihin?
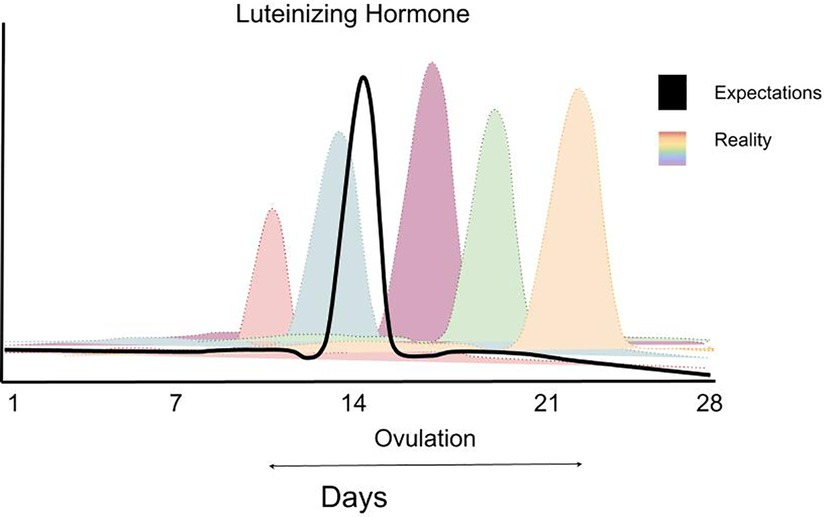
Kuva 2: Kuukautiskierron vaiheet ja hormonipitoisuuksien muutokset. Ensimmäinen kierron vaiheista on follikkelivaihe, toinen vaihe on ovulaatio, eli munasolun irtoaminen ja kolmas vaihe luteaalivaihe. Estradiolipitoisuus saavuttaa yleensä huippupitoisuutensa juuri ennen ovulaation alkua.
Subjektiivisesti kuukautiskierto vaikuttaa suorituskykyyn, muttei objektiivisesti?
Vaikka urheilijat itse kokevat kuukautiskierron vaikuttavan suorituksiinsa, objektiiviset tutkimustulokset eivät aina tue tätä käsitystä. Esimerkiksi 241 urheilijan tutkimuksessa ei havaittu suorituskyvyn eroja eri kierron vaiheissa (6).
Hormonitoiminnalla on selitetty suorituskyvyn vaihtelua
Estrogeenihormonia on pidetty keskeisenä tekijänä follikulaarisen vaiheen suorituskyvyn nousussa, kun taas keltarauhashormoni eli progestiini yhdistetään luteaalivaiheen heikentyneeseen suorituskykyyn (7). Progestiini nostaa kehon lämpötilaa, mikä voi haitata aineenvaihduntaa ja aiheuttaa lisärasitusta sydän- ja verisuonijärjestelmälle (8). Estrogeeni puolestaan tehostaa glukoosin ottoa lihakseen supistuksen jälkeen, kun taas progestiini saattaa estää tätä prosessia (9). Lisäksi estrogeeni edistää vapaiden rasvahappojen käyttöä energialähteenä, mikä säästää lihasten glykogeenivarastoja liikunnan aikana (7).
Lyhyesti yksi estrogeenin ilmentymismuodon, estradiolin, vaikutus suorituskykyyn on merkittävä. Estradioli voi edistää lihaskudoksen rakentumista, voimantuottoa ja aineenvaihduntaa säästämällä lihasten energiavarastoja sekä lisäämällä hiilihydraattien käyttöä. Sillä on myös antioksidanttisia ja tulehdusta vähentäviä vaikutuksia, jotka suojaavat lihaskudosta vaurioilta. Lisäksi estradioli voi parantaa hermoston toimintaa ja lihasten tahdonalaista aktivointia.
Olisi loogista olettaa, että suorituskyky vaihtelee kuukautiskierron vaiheiden mukaan, erityisesti silloin, kun estrogeenitasot ovat alhaalla – esimerkiksi myöhäisessä luteaalivaiheessa tai aikaisessa follikulaarivaiheessa. Kuitenkin suorituskyvyn on havaittu olevan yllättävän vakaa kierron eri vaiheissa.
Harjoittelun ohjelmointi kuukautiskierron mukaan
Harjoittelun ohjelmointia kuukautiskierron mukaan tukee vain muutama tutkimus. Näistä tarkemmin vain kaksi artikkelia on raportoinut, että harjoittelun keskittäminen follikulaariseen vaiheeseen, jolloin harjoitusfrekvenssi ja -volyymi ovat korkeampia, tuottaa parempia tuloksia verrattuna luteaalivaiheen painottamiseen tai tasaisesti kierron eri vaiheisiin jaettuun harjoitteluun (24, 25). Laajemmin tarkasteltuna tulokset eivät kuitenkaan ole yksiselitteisiä. Uudessa katsauksessa (26), jossa huomioitiin kaikki saatavilla oleva data, tulokset vaihtelivat merkittävästi. Suuri osa tutkimuksista oli metodologisesti heikkotasoisia, eikä niiden perusteella voida tehdä luotettavia johtopäätöksiä harjoittelun ohjelmoinnin hyödyistä kuukautiskierron mukaan. Nykyisen tiedon perusteella kuukautiskierron mukaisella harjoittelulla ei ole pitkän aikavälin vaikutusta voiman, suorituskyvyn tai lihaskasvun kehitykseen.
Mutta miksi tässä on niin räikeä ero subjektiivisesti koetun ja objektiivisesti mitattujen tekijöiden välillä?
Kuva 3: Hypotalamus-aivolisäke-munasarja-akselin normaali toiminta. Lähde: Luiro-Helve ja Holopainen. Lääketieteellinen Aikakauskirja Duodecim 2023;139(5):359-367
Metodologia ei huomioi yksilöllisiä eroja
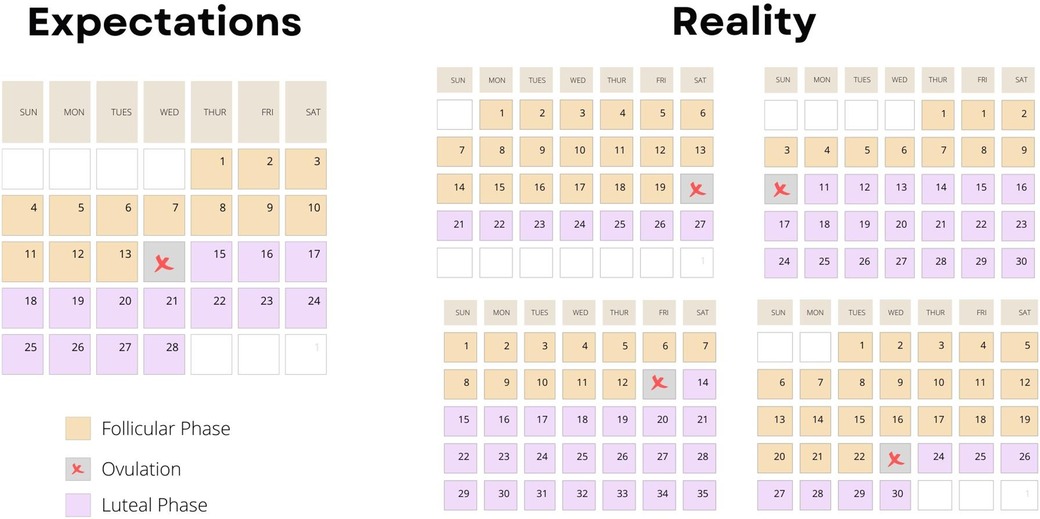
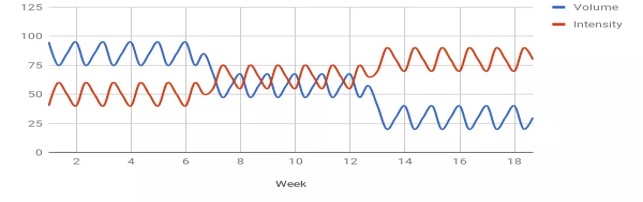
Kuukautiskierron vaiheen määrittämisen haasteet korostuvat tutkimusten ristiriitaisissa tuloksissa. Follikulaarisen vaiheen kesto voi vaihdella 10–22 päivän välillä ja luteaalivaiheen 7–17 päivän välillä (27) (katso kuva 4). Jo vuonna 1926 Ludvig Fränkel totesi, että kuukautiskierron ainoa säännöllisyys on epäsäännöllisyys (28). Tästä huolimatta naisten kuukautiskierron oletetaan yhä olevan tasainen 28 päivän sykli, jossa ovulaatio tapahtuu kolmantenatoista päivänä (kuva 5). Esimerkiksi luteinisoivan hormonin monitorointi, yksi yleisimmin käytetyistä kierron vaiheiden määrittämismenetelmistä, voi osoittaa ovulaation yhden päivän tai 2–6 päivän ajalta riippuen yksilöstä (29).
Kuva 4: Kaaviokuva, joka havainnollistaa luteinisoivan hormonin piikin vaihtelua naisilla ja korostaa sitä, kuinka epätarkoituksenmukaista on suunnitella suurempaa harjoitusvolyymia kuukautiskierron tietylle vaiheelle. Tiedot ovat teoreettisia, mutta perustuvat havaintoihin, jotka osoittavat LH-piikin huomattavaa vaihtelua naisilla. Lähde: 26.
Sarwar et al. (1996) (10) havaitsivat, että voima kasvaa ovulaation aikana, mikä yhdistettiin estrogeenitasojen nousuun. Näitä tuloksia ei kuitenkaan ole pystytty toistamaan muissa tutkimuksissa, kuten penkkipunnerruksen maksimitoistoissa tai aerobisessa suorituskyvyssä (11). Tuoreessa meta-analyysissa (12) havaittiin vähäinen suorituskyvyn heikkeneminen varhaisessa follikulaarivaiheessa, mutta tutkimusten vähäinen määrä ja laatu tekevät johtopäätöksistä epävarmoja. Toinen systemaattinen katsaus (13), jossa tutkittiin 314 eliittiurheilijaa, ei löytänyt todisteita kuukautiskierron vaikutuksesta suorituskykyyn. Toisaalta vastakkaisiakin tuloksia on, jossa huomattiin että varhainen follikulaarivaihe on epäedullinen kaikille voimatyypeille. Isometrinen voima on huipussaan myöhäisessä follikulaarivaiheessa, kun taas isokineettinen voima saavuttaa huippunsa ovulaation aikana. Dynaaminen voima on optimaalisimmillaan myöhäisessä follikulaarivaiheessa (30).
On selvää, että itsearvioidun kuukautiskierron vaikutusten ja todellisten fysiologisten suorituskykyerojen välillä on eroja. Tarvitaan lisää korkealaatuista tutkimusdataa, jotta voidaan ymmärtää kuukautiskierron vaiheen ja suorituskyvyn monimutkaista vuorovaikutusta.
Kuva 5: Havainnollistava kuva, joka osoittaa kuinka epätarkoituksenmukaista on suunnitella suurempaa harjoitusvolyymia kuukautiskierron tietylle vaiheelle, jos oletetaan ovulaation ajankohdan tapahtuvan aina samassa vaiheessa kuukautta. Lähde: 26.
Mutta onko kyseessä kuukautiskierron aiheuttama suorituskyvyn lasku vai laiskaa valmennusta?
Kuukautiskierron häiriöt voivat johtua useista tekijöistä, joista yksi merkittävimmistä on krooninen energiavaje. Tämä voi johtaa hypotalamisesta amenorreaan, jossa kuukautiset jäävät väliin hypotalamuksen toimintahäiriön seurauksena (3). Hypotalaminen amenorrea on huomattavan yleinen erityisesti kestävyysjuoksijoilla, joista jopa 65 % kärsii tästä tilasta, sekä balettitanssijoilla, joiden kohdalla esiintyvyys on jopa 79 %. Yleisväestössä vastaava esiintyvyys on vain noin 5 % (4, 5). Tämä viittaa siihen, että kuukautiskiertoon liittyvät suorituskyvyn haasteet eivät välttämättä johdu pelkästään kierron fysiologisista vaiheista, vaan taustalla voi olla muita tekijöitä, kuten energiatasapainon häiriöt, jotka ovat hallittavissa asianmukaisilla toimenpiteillä ja tuella. Toinen merkittävä tekijä on runsaat kuukautisvuodot, jotka voivat altistaa raudan puutokselle. Raudalla on tärkeä rooli elimistön hapenkuljetuksessa ja sen puute heikentää suorituskykyä.
Onko ehkäisytavalla väliä suorituskyvyn näkökulmasta?
Pyrkimyksenä lieventää kehon sisäisten sukupuolihormonien vaihtelun mahdollisia fysiologisia vaikutuksia – tai ainakin vähentää heikomman suorituskyvyn kokemusta kuukautiskierron aikana – urheilijat saattavat käyttää hormonaalista ehkäisyä kierron hallitsemiseksi. Hormonaalista ehkäisyä hyödynnetään usein kuukautisten alkamisen viivästyttämiseen tietyn kilpailupäivän yli, kierron oireiden lievittämiseen tai esimerkiksi runsaan kuukautisvuodon aiheuttaman raudanpuuteanemian ehkäisyyn. Joissakin tapauksissa urheilijat voivat trisyklisellä lähestymistavalla välttää kuukautisten tyhjennysvuodon koko kilpailukauden ajan.
Hormonaalisen ehkäisyn vaikutukset suorituskykyyn
Vaikka hormonaalista ehkäisyä käytetään urheilijoiden keskuudessa laajasti, tutkimukset eivät ole osoittaneet sen parantavan suorituskykyä merkittävästi. Jonesin ym. (31) kyselytutkimuksen mukaan suorituskyvyn koettu parantuminen oli vähäistä hormonaalista ehkäisyä käyttävillä urheilijoilla. Lisäksi tuoreessa meta-analyysissa (14) todettiin, että hormonaalisen ehkäisyn käyttö voi aiheuttaa pienen mutta todennäköisesti negatiivisen vaikutuksen urheilusuoritukseen. Esimerkiksi ehkäisypillereiden käytöllä ei ole havaittu positiivisia vaikutuksia lihasvoimaan verrattuna ei-hormonaaliseen ehkäisyyn (15), ja se voi jopa laskea maksimaalista hapenottokykyä (16).
Näiden tulosten perusteella hormonaalisen ehkäisyn käyttöä ei voida suositella suorituskyvyn optimoimisen näkökulmasta. Toisaalta ehkäisyn käytöllä voi olla tärkeitä sosiaalisia ja henkisiä hyötyjä, jotka ovat merkityksellisiä suorituskyvyn ulkopuolella. Esimerkiksi Jonesin (31) tutkimuksessa 81 % ei-hormonaalista ehkäisyä käyttävistä raportoi kipeitä kuukautiskipuja, kun taas hormonaalista ehkäisyä käyttävistä vain 53 % raportoi samanlaisia oireita. Lisäksi jotkut tutkimukset ovat osoittaneet, että hormonaaliset ehkäisymenetelmät eivät välttämättä vaikuta suorituskykyyn merkittävästi (17).
Minipillerit ja urheilijat
Pelkkää progestiinia, eli keltarauhashormonia, sisältävät ehkäisyvalmisteet, kuten minipillerit ja pitkävaikutteiset menetelmät (esim. kapselit ja kierukat), ovat kasvattaneet suosiotaan viime vuosina. Näillä valmisteilla on monia etuja, kuten se, että ne eivät lisää veritulppariskiä. Mutta progestiinipohjaisten menetelmien mahdollinen negatiivinen vaikutus suorituskykyyn pitkällä aikavälillä on selkeästi osoitettu ja huippu-urheilussa pienikin myönnytys kehittymisen suhteen voi olla ratkaiseva. Keltarauhashormonilla on esimerkiksi monilta osin estradiolin toimintaa hillitsevä vaikutus. Tällä hetkellä ei kuitenkaan ole riittävästi tutkimustietoa suositella mitään tiettyä ehkäisymenetelmää yleisesti huippu-urheilijoille. Siksi urheilijoiden kannattaa keskustella aiheesta paitsi terveydenhuollon ammattilaisten myös valmentajiensa kanssa löytääkseen itselleen parhaiten sopivan ratkaisun.
Yhteenveto
Kuukautiskierron mukainen harjoittelun ohjelmointi ei toimi, eikä sitä kannata käyttää. Tulosten puute johtuu luultavasti metodologisista ongelmista ja kuukautiskiertojen eri vaiheiden määrittelyn vaikeudesta. Ennen kuin lisää kattavaa dataa saadaan, niin en hukkaisi aikaa tähän. Kuukautiskierron mukainen harjoittelun ohjelmointi ei ole näyttöön perustuva lähestymistapa nykyisellä tietämyksellämme. Laadukkaan näytön puuttuessa kuukautiskiertoon perustuvien harjoitusohjelmien räätälöimisessä valmentajien ja urheilijoiden tulisi rakentaa ohjelmointi yksilön mukaan. Kuukautiskierron vaikutus voi olla huomioitava tekijä ohjelmaa suunniteltaessa, samoin kuin monet muut tutut tekijät, kuten: ravitsemus, väsymys, unen laatu, stressi, loukkaantumiset, motivaatio ja muut harjoittelun sisäiset muuttujat. Myös kuukautisoireet voivat vaikuttaa joidenkin naisten harjoitussuoritukseen merkitsevästi. Tämän takia urheilijan omaa tuntemusta kannattaa kuunnella herkällä korvalla.
Lähteet:
1. Costello JT, Bieuzen F, Bleakley CM. Where are all the female participants in sports and exercise medicine research? Eur J Sport Sci. (2014) 14(8):847–51. doi: 10.1080/17461391.2014.911354
2. Cowley ES, Olenick AA, McNulty KL, Ross EZ. “Invisible sportswomen”: the sex data gap in sport and exercise science research. Women Sport Physical Activity J. (2021) 29(2):146–51. doi: 10.1123/wspaj.2021-0028
3. Jones BP, L’Heveder A, Saso S, Yazbek J, Smith JR, Dooley M. Sports gynaecology. Obstet Gynaecol. (2019) 21(2):85–94. doi: 10.1111/tog.12557
4. Dusek T. Influence of high intensity training on menstrual cycle disorders in 56 athletes. Croat Med J. (2001) 42:79–82.11172662
5. Abraham SF, Beumont PJV, Fraser IS, Llewellyn-Jones D. Body weight, exercise and menstrual status among ballet dancers in training. Br J Obstet Gynaecol. (1982) 89:507–10. doi: 10.1111/j.1471-0528.1982.tb03649.x
6. Kishali NF, Imamoglu O, Katkat D, Atan T, Akyol P. Effects of menstrual cycle on sports performance. Int J Neurosci. (2006) 116(12):1549–63. doi: 10.1080/00207450600675217
7. Lebrun CM, Joyce S, Constantini NW. Effects of female reproductive hormones on sports performance. In: Constantini N, Hackney A, editors. Endocrinology of Physical Activity and Sport. Contemporary Endocrinology. Totowa: Humana Press (2013). p. 267–302.
8. Janse de Jonge XA. Effects of the menstrual cycle on exercise performance. Sports Med. (2003) 33(11):833–51. doi: 10.2165/00007256-200333110-00004
9. Oosthuyse T, Bosch AN. The effect of the menstrual cycle on exercise metabolism: implications for exercise performance in eumenorrhoeic women. Sports Med. (2010) 40(3):207–27. doi: 10.2165/11317090-000000000-00000
10. Sarwar R, Niclos BB, Rutherford OM. Changes in muscle strength, relaxation rate and fatiguability during the human menstrual cycle. J Physiol. (1996) 493(Pt 1):267–72. doi: 10.1113/jphysiol.1996.sp021381
11. Quadagno D, Faquin L, Lim G-N, Kuminka W, Moffatt R. The menstrual cycle: does it affect athletic performance? Phys Sportsmed. (1991) 19(3):121–4. doi: 10.1080/00913847.1991.11702172
12. McNulty KL, Elliott-Sale KJ, Dolan E, Swinton PA, Ansdell P, Goodall S, et al. The effects of menstrual cycle phase on exercise performance in eumenorrheic women: a systematic review and meta-analysis. Sports Med. (2020) 50(10):1813–27. doi: 10.1007/s40279-020-01319-3
13. Meignié A, Duclos M, Carling C, Orhant E, Provost P, Toussaint JF, et al. The effects of menstrual cycle phase on elite athlete performance: a critical and systematic review. Front Physiol. (2021) 12:654585. doi: 10.3389/fphys.2021.654585
14. Elliott-Sale KJ, McNulty KL, Ansdell P, Goodall S, Hicks KM, Thomas K, et al. The effects of oral contraceptives on exercise performance in women: a systematic review and meta-analysis. Sports Med. (2020) 50(10):1785–812. doi: 10.1007/s40279-020-01317-5
15. Nichols AW, Hetzler RK, Villanueva RJ, Stickley CD, Kimura IF. Effects of combination oral contraceptives on strength development in women athletes. J Strength Cond Res. (2008) 22(5):1625–32. doi: 10.1519/JSC.0b013e31817ae1f3
17. Elliott-Sale, K. J., Mcnulty, K. L., Ansdell, P., Goodall, S., Hicks, K. M., Thomas, K., Swinton, P. A., & Dolan, E. (2020). The effects of oral contraceptives on exercise performance in women: A systematic review and meta-analysis. Sports Medicine (Auckland, N.Z.), 50(10), 1785–1812.
18. Chidi-Ogbolu N BK. Effect of estrogen on musculoskeletal performance and injury risk. Front Physiol. (2019) 9:1834. 10.3389/fphys.2018.01834
19. Collins BC, Arpke RW, Larson AA, Baumann CW, Xie N, Cabelka CA, et al. Estrogen regulates the satellite cell compartment in females. Cell Rep. (2019) 28:368–81.
20. Ikeda K, Horie-Inoue K, Inoue S. Functions of estrogen and estrogen receptor signaling on skeletal muscle. J Steroid Biochem Mol Biol. (2019) 191:105375. 10.1016/j.jsbmb.2019.105375
21. Chidi-Ogbolu N BK. Effect of estrogen on musculoskeletal performance and injury risk. Front Physiol. (2019) 9:1834. 10.3389/fphys.2018.01834
22. Collins BC, Arpke RW, Larson AA, Baumann CW, Xie N, Cabelka CA, et al. Estrogen regulates the satellite cell compartment in females. Cell Rep. (2019) 28:368–81.
23. Ikeda K, Horie-Inoue K, Inoue S. Functions of estrogen and estrogen receptor signaling on skeletal muscle. J Steroid Biochem Mol Biol. (2019) 191:105375. 10.1016/j.jsbmb.2019.105375
24. Sung E, Han A, Hinrichs T, Vorgerd M, Manchado C, Platen P. Effects of follicular versus luteal phase-based strength training in young women. SpringerPlus. (2014) 3:668. 10.1186/2193-1801-3-668
25. Wikström-Frisén L, Boraxbekk CJ, Henriksson-Larsén K. Effects on power, strength and lean body mass of menstrual/oral contraceptive cycle-based resistance training. J Sports Med Phys Fitness. (2017) 57:43–52. 10.23736/S0022-4707.16.05848-5
26. Colenso-Semple LM, D’Souza AC, Elliott-Sale KJ, Phillips SM. Current evidence shows no influence of women’s menstrual cycle phase on acute strength performance or adaptations to resistance exercise training. Front Sports Act Living. 2023 Mar 23;5:1054542. doi: 10.3389/fspor.2023.1054542. PMID: 37033884; PMCID: PMC10076834.
27. Shea A, Vitzthum V. The extent and causes of natural variation in menstrual cycles: integrating empirically-based models of ovarian cycling into research on women’s Health. Drug Discov Today Dis Models. (2020) 32:41–9. 10.1016/j.ddmod.2020.11.002¨
28. Vitzthum V. Field methods and strategies for assessing female reproductive functioning. Am J Hum Biol. (2021) 33:e23513. 10.1002/ajhb.23513
29. Su HW, Yi YC, Wei TY, Chang TC, Cheng CM. Detection of ovulation, a review of currently available methods. Bioeng Transl Med. (2017) 2(3):238–46. 10.1002/btm2.10058
30. Niering M, Wolf-Belala N, Seifert J, Tovar O, Coldewey J, Kuranda J, Muehlbauer T. The Influence of Menstrual Cycle Phases on Maximal Strength Performance in Healthy Female Adults: A Systematic Review with Meta-Analysis. Sports. 2024; 12(1):31. https://doi.org/10.3390/sports12010031
31. Jones, B. P., L’Heveder, A., Bishop, C., Kasaven, L., Saso, S., Davies, S., Chakraverty, R., Brown, J., & Pollock, N. (2024). Menstrual cycles and the impact upon performance in elite British track and field athletes: a longitudinal study. Frontiers in sports and active living, 6, 1296189. https://doi.org/10.3389/fspor.2024.1296189
Lihaskasvua käsittelevä artikkelisarja lähenee loppua ja on viimeisen osan vuoro. Tässä artikkelissa käsitellään miten eri yksilöt adaptoituvat voimaharjoitteluun ja miten harjoittelu kannattaisi ohjelmoida, jotta kehitys olisi kaikkein optimaalisinta.
Genetiikan vaikutus lihaskasvuun
Geenit vaikuttavat suuresti henkilön kehittymiseen. Teoreettisesti voidaan ajatella maksimaalisen lihaskasvun rajojen olevan olemassa. Nämä määrittyvät genotyypin, eli geneettisen informaation, ja näiden geenien ilmentymisen yksilössä mukaan. Geenit asettavat teoreettisesti meille tietyt reunaehdot, miten reagoimme erilaisiin tilanteisiin. Geenien takia jotkut yksilöt kehittyvät nopeammin ja jotkut yksilöt hitaammin. Kuiinka paljon vaihtelua voi sitten eri henkilöiden välillä olla?
KUVA 1: Tarkistuslista, jos epäilet salimarkun tai salijonnan käyttävän kehon ulkopuolisia hormoneita.
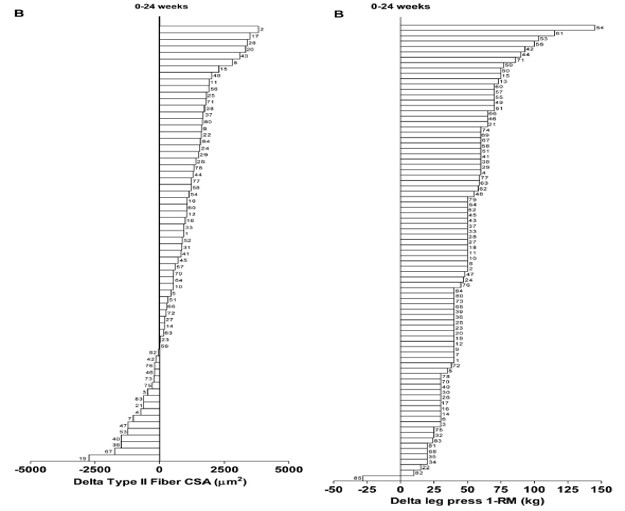
Yksilölliset vasteet harjoitteluun vaihtelevat hurjasti ihmisillä. Tämä tarkoittaa sitä, että jos kaksi samaa ihmistä tekee täysin saman ohjelman niin tuskin koskaan he kehittyvät täsmälleen saman verran. Esimerkiksi samanlaisella harjoittelulla lihaksen kasvu voi vaihdella jopa -11 % - + 30 % välillä ja voima taas - 8 % - + 60 % välillä (Ahtiainen et al., 2016). Ei-respondoivia ihmisiä, eli niitä, jotka eivät reagoi mitenkään tutkimusten voimaharjoitteluun on keskimäärin 25% koehenkilöistä (Bamman et al., 2007). Toisaalta nykyään ajatellaan, että erilainen harjoittelu, ravinto, lepo tai joku muu muuttuja olisi voinut vaikuttaa yksilöllisiin vasteisiin. Eli tälläiset ei-respondoivat koehenkilöt johtuvat pääosin huonosta tutkimusasetelmasta, elämäntilanteesta tai muusta tekijästä, mikä estää kehityksen. Jotkut ei-respondoivista ihmisistä tarvitsevat vain enemmän ja pidempää harjoittelua verrattuna toisiin.
KUVA 2: Oikeassa kaaviossa nähdään, että lähes kaikki koehenkilöt saivat erään tutkimuksen aikana lisää voimaa, mutta vasemmalla huomataan, että joidenkin koehenkilöiden lihaskoko jopa pieneni tutkimuksen aikana. (Churward-Venne et al., 2015).
Mikä näitä eroja sitten selittää? Pescatello ja kummanit (2013) pystyivät tunnistamaan 17 geeniä, joiden uskotaan selittävän osan henkilöiden välisistä eroista. Näistä tärkeimpänä hypertrofian kannalta pidetään tällä hetkellä BMP2-geeniä (Devaney et al., 2009). Ja myös MGF:ää pidetään erittäin tärkeänä (Bamman et al., 2007).
Lisäksi satelliittisolut lisääntyvät ja luovuttavat tumia tehokkaammin hyvin respondoituvilla ihmisillä kuin verrataan ei-respondoituviin ihmisiin (Petrella et al., 2008). Myös lihaksen morfologia vaikuttaa suuresti lihaskasvuun. Mitä enemmän lihassoluja henkilöllä on, niin sen helpommin lihas kasvaa. Lihassolujen määrä on aika vakio ja hyperplasiaa (lihassolujen lisääntymistä) ei tapahdu enää 24 viikon jälkeen syntymästä. Lihassolujen määrä onkin pitkälti geenien määrittämä.
Jos vertaillaan mieskehonrakentajia ja tavallisia kadun tallaajia niin kehonrakentajilla on enemmän lihassoluja hauiksissa ja nämä solut olivat myös isompia verrattuna tavallisiin tallaajiin (MacDougall et al., 1984). Joten niillä, jotka päätyivät kehonrakentajiksi oli jo enemmän lihassoluja kuin muilla normaaleilla ihmisillä. Tämä on yksi syy, miksi kehonrakentajat reagoivat paremmin voimaharjoitteluun.
Nopeiden lihassolujen määrä kasvatettavassa lihaksessa on tärkeä tekijä sen suhteen miten voimaharjoittelu vaikuttaa lihakseen. Tyypin II - lihassolut kasvavat noin 50% enemmän verrattua tyypin I -lihassoluihin. Toisaalta tässäkin suhteessa on hurjasti henkilöiden välistä vaihtelua (Kosek et al., 2006). Tutkimuksia, jotka selittävät fysiologisia mekanismeja yksilöllisessä voimaharjoittelussa on saatavilla erittäin niukasti, mutta erot johtuvat yleisesti perimästä, geenien ilmenemisessä, soluviestinnässä, hormonireseptoreiden toiminnassa ja satelliittisoluissa. Nostan vielä esille ravitsemuksen, jota harvoin tutkimuksissa kontrolloidaan ja sillä on valtava merkitys lihaskasvun mahdollistamisessa.
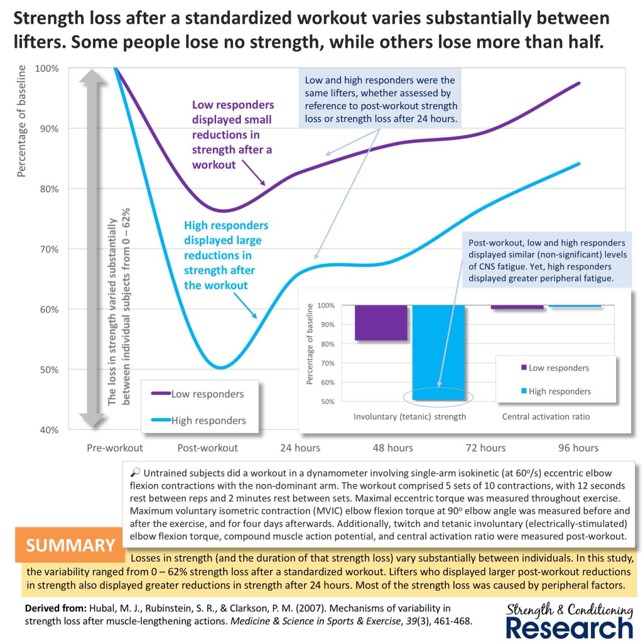
KUVA 3: Näyttäisi myös siltä, että hyvin voimaharjoitteluun reagoivat ihmiset saavat itsestään irti paljon harjoituksen aikana. Heillä harjoituksen jälkeen voima laskee enemmän kuin huonosti respondoituvilla ihmisillä. Tämä tarkoittaa sitä, että he ovat saaneet enemmän väsymystä aikaiseksi harjoituksen aikana.
Genetiikalla on siis kieltämättä iso rooli henkilön lihaskasvussa. Voit kiittää vanhempiasi näistä lahjoista, mutta muuten asiaan ei kannata kiinnittää sen enempää huomiota. Jokainen voi ja pystyy kehittymään, vaikka tutkimuksissa usein löydetään ei-respontoivia henkilöitä harjoitteluun niin tähän saattaa olla syynä vääränlainen tutkimuksenasettelu, harjoittelu tai ohjelmointi. Yksilöllinen harjoittelu ja ohjelma mahdollistavat jokaisen kehittymisen. Heikosti respondoivien pitää joskus reenata jopa paljon vähemmän ja harvemmin kehittyäkseen. Tai kevyemmillä kuormilla. Myös harjoitustausta vaikuttaa suuresti henkilön kykyyn adaptoitua. Aloittelevilla ihmisillä lihaskasvu voi olla jopa 28% ja korkeimmillaan 58% 16 viikossa (Stewart & Rittweger, 2006), kun taas kehonrakentajat kehittyivät viidessä kuukaudessa “vain” 3-7% (Alway et al., 1992). Ohjelmointi ja ohjelman muuttujien manipulointi korostuu erityisesti, kun harjoitustaustaa kertyy enemmän. Kevennetyt jaksot voivat auttaa taas respontoimaan paremmin stimulukseen (Ogasawara et al., 2013).
Ei-respondereilla ei siis ole mielenvika pääsään vaan se on ihan oikea asia. Heidän tulee erityisesti keskittyä ravintoon ja oikeanlaiseen harjoitteluun. Huonosti respondoivat saattavat olla vain hitaammin respondoivia ja tarvitsevat enemmän harjoitusvolyymia kehittyäkseen. Nimi on vähän harhaanjohtava sillä jokainen kehittyy oikeanlaisella harjoittelulla. Saatat vain olla heikommin ja hitaammin respondoituva henkilö verrattuna salikaveriisi. Lisäksi vielä useissa tutkimuksissa kehitys on 12 viikossa esim. non respondereilla “vain” 5%. Tämä tulos luokitellaan tutkimuksissa hitaasti reaspondoivaksi, mutta jos kehitys jatkuisi koko vuoden verran samanlaisena se vastaisi alle vuodessa esimerkiksi 100kg penkissä siirtymistä 121,6 kiloon. Kuinka moni olisi tyytyväinen tuohon kehitykseen vuodessa? Minä ainakin olisin. Joten nimi on hiukan harhaanjohtava ja yleisesti ongelmat johtuvat tutkimusasetelmasta, sekä heterogeenisestä tutkimusjoukosta.
Neuvostoliitossa
periodisaation (ja ylipäätään liikuntatieteen) tutkimus- ja kehitystyö oli
tuolloin huomattavan laajaa ja systemaattista. Tämän tutkimustyön tuloksena
syntyi klassinen periodisaatiomalli. Muissakin maissa toki käytettiin ja
kehitettiin samantyylistä periodisaatiota, tosin pienemmässä mittakaavassa.
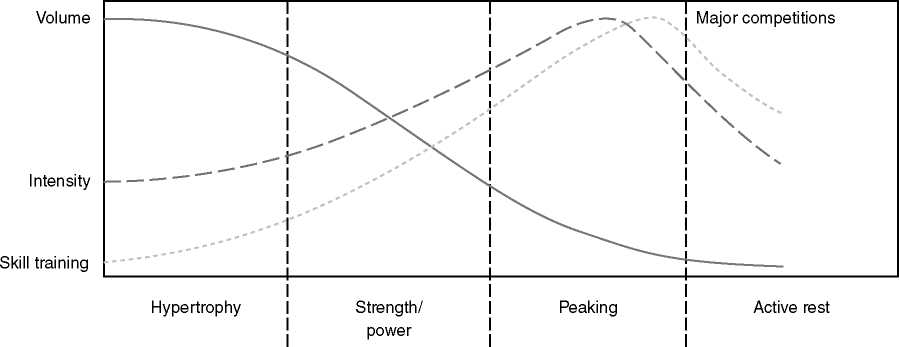
KUVA 4: Klassinen periodisaatiomalli, missä harjoitusvolyymi on aluksi korkealla ja vähenee pääkilpailun lähestyessä. Samalla harjoittelun intensiteetti nousee.
Periodisaatio
on erittäin vähän tutkittu aihealue. Johtuen luultavasti siitä, että
periodisaatio ei ole mikään selkeästi käsitettävä kokonaisuus. Erillaisia
malleja voidaan esittää, mutta näidenkin sisällä voi olla loputtamattomasti
erillaisia muutujia, joita voi siirrellä ja muutella. Periodisaatio perustuu
Selyen yleiseeen adaptaatio syndrooma - teoriaan (GAS - general adaptation
syndrome). Se perustuu kehon reaktioihin, jotka ovat hälytys, vastustus ja
uupuminen. (Selye, 1950). Teoriaa on nykypäivinä kritisoitu
yksinkertaistamisesta ja siitä, että se ei huomioi esimerkiksi psykologisia,
teknisiä ja ravinnollisia seikkoja. Selyen teorian mukaan harjoittelua
pidetään stressitekijänä, johon voidaan soveltaa GAS - teoriaa. Harjoittelun
tulee olla progressiivisesti haastavaa, jotta tasannevaihetta ei tule
harjoittelussa ja toisaalta harjoittelu ei saa olla ylikuluttavaa.
Periodisaatiomalleista kolmea on tutkittu lihashypertrofian kannalta: perinteinen lineaarinen malli, ei-lineaarinen ja käänteinen periodisaatio.
Perinteisessä
lineaarisessa mallissa on vähintään makrosykli, mesosykli ja mikrosykli
(esimerkiksi vuosi, peruskuntokausi 8 vko, yksi harjoitusviikko). Malli etenee
korkeasta voluumista ja matalasta intensiteetistä kohti korkeaan
intensiteettiin ja matalaan voluumiin. Esimerkiksi rakenteellinen
jakso/hypertrofinen jakso, jossa voluumi nousee ja intensiteetti pysyy 60-75%
välillä, seuraavaksi voi olla perusvoimakausi intensiteetin ollessa 80-90%
välillä ja voluumin vähetessä. Viimeinen mesosykli olisi intensiteetiltään
suurin 90-100%, mutta voluumiltaan pienin.
Lihaskasvun osalta aihetta on tutkittu jonkin verran, mutta itse pidän tutkimuksia metodologisilta osuuksilta ontuvilta, joten johtopäätöksiä ei tule vetää vielä näiden tutkimusten pohjalta. Stone ja kumppanit (1981) tutkivat periodisaatioidun ohjelman ja ei-periodisoidun ohjelman eroja, ja huomasivat hypertrofian (mitattu veden alaispunnituksella) eroavan tilastollisesti merkitsevästi ei-periodisoidusta ohjelmoinnista. Baker:in tutkimusporukka taa (1994) ei löytänyt eroja periodisatioidun ja ei-periodisaatiouidun välille (ihopoimumittaus). Kun taas Monteiro ja kumppanit (2009) löysivät eron (ihopoimu, ei tilastollisesti merkittävä) periodisoidun harjoittelun hyväksi.
Voiman osalta periodisoitu harjoittelu tuottaa kiistatta parempia tuloksia kuin ei periodisoitu harjoittelu (Ahmadizad et al., 2014;, Monteiro et al., 2009,Willoughby, 1993). Voisi ajatellakin, että koska voima nousee paremmin periodisoidulla harjoittelulla, johtaisi se mekaanisen kuormituksen johdosta myös suurempiin hypertrofisiin adaptaatioihin ajan kanssa, mutta tätä ei ole vielä todistettu.
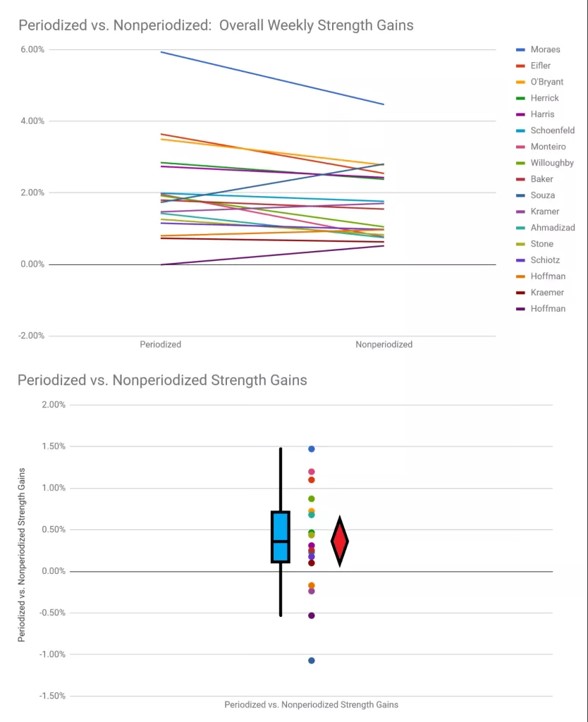
KUVA 5: Periodisoitu harjoittelu tuottaa keskimäärin 21,78 – 23,62% voiman nousun, kun taas ei periodisoitu harjoittelu johtaa vain 18,90 – 19,10% keskimääräiseen voiman nousuun. Viikkotasolla samat luvut 1,96 % - 2,05 % verrattuna ei periodisoituun 1,59 – 1,70%. (Nuckols, 2018.)
Ei-lineaarinen periodisaatio tai aaltoileva periodisaatio on nykyään yksi suosituimmista suuntauksista. Sitä on käytetty jo kauan, mutta Poliquin (1988) oli ensimmäinen, joka esitteli sen tiedeyhteisössä. Hänen mukaansa lineaarisen ohjelmoinnin ongelmat ovat liian suuri progressiivinen painon lisäys liian lyhyessä ajassa ja alussa saavutettujen hypertrofisten adaptaatioiden menetys muissa mesosykleissä, koska voluumi laskee liikaa.
Aaltoilevassa periodisaatiossa vaihdellaan voluumia ja intensiteettiä kuinkas muutenkaan kuin aaltoilevaan tyyliin. Mallia on kehitetty eteenpäin ja tällä hetkellä suosituin malli taitaa olla päivittäin vaihtuva periodisaatio (daily undulating periodization - DUP). Tässä mallissa on viikon sisällä maksimivoima, perusvoima ja hypertrofinen harjoitus. Aihetta on tutkittu suhteellisen laajasti. Ja vain yksi tutkimus sai tilastollisesti merkitsevän eron lineaarisen ja ei-lineaarisen mallin välille. Tässä tutkimuksessa ei-lineaarinen malli oli selvästi parempi lisäämään kyynärpään koukistajien ja ojentajien tiheyttä (Simao et al., 2012). Muuten muissa tutkimuksissa ei ole saatu eroja mallien välille lihaskasvun osalta, mikä osoittaa kummankin mallin olevan yhtä toimiva lihaskasvun osalta (Baker et al., 1994; Davitt et al., 2014; , Harries et al., 2015; Monteiro et al., 2009, Prestes et al., 2009, Simao et al., 2012). Toisaalta tutkimusten kesto on ollut korkeimmillaan vain 12 viikkoa ja kun puhutaan periodisaatiosta yleisesti urheilijoiden osalta puhutaan vuosista.
Itse suosin valmennettavillani enemmän hybridimallia, joka on versio aaltoilevasta mallista. Tässä mallissa harjoitusohjelma on laadittu blokkeihin ja näiden blokkien sisällä tapahtuu aaltoilua. Blokit toisaalta etenevät lineaarisesti. Viikon sisällä siinä tehdään maksimivoimasarjoja ja perusvoima sarjoja samassa harjoituksessa järkevästi ohjelmoiden. Pitkillä perusvoimakausilla saattaa hävitä liikaa maksimivoimaa ja pitkillä maksimivoima kausilla saattaa hävitä liikaa lihasta tai perusvoimaa, joten järkevä hybridiohjelmointi poistaa nämä ongelmat.
KUVA 6: Aaltoileva periodisaatio, missä volyymi ja intensiteetti aaltoilevat mahdollistaen intensiteetin ylläpidon peruskuntokaudella ja volyymin ylläpidon kisakaudella.
Käänteistä periodisaatiota on ehdotettu lihaskasvuun tähtääville tärkeimmäksi ohjelmaksi. Lineaarinen malli tiputtaa jatkuvasti voluumia kohti kauden päätähtäintä ja kun tiedetään voluumin olevan elintärkeä hypertrofian kannalta voisi ajatella lineaarisen mallin olevan huono lihaskasvun kannalta. Käänteisessä mallissa voluuumi on korkeimmillaan kauden päätähtäimen kohdalla. Aihe on vielä tutkimuskentällä tuore ja vain tietääkseni Prestes ja kumppanit (2009) ovat sitä tutkineet huonoin tuloksin käänteisen mallin puolesta. Lineaarinen malli oli huomattavasti parempi kuin käänteinen malli. Toisaalta mittauksissa käytettiin ihopoimupihtejä, jotka ovat metodisesti yhtä tyhjän kanssa.
Tiettyä suunnitelmallisuutta harjoitteluun tarvitaan, koska muuten harjoittelusta puuttuu variaatio, progressiivisuus ja tavoitteet. Lihaskasvun osalta yksi tärkeimmistä periodisaation tuomista asioista on kevennetyt jaksot. Keho ottaa paremmin vastaan harjoittelua kuin välillä tiputtaa voluumia ja antaa kropan palautua. Lisäksi on tärkeä kerätä palautetta periodisaation toimivuudesta ja suunnitelman muokkaaminen sen mukaan auttavat kehittämään ohjelmointia.
Kiitos lukijalle, jos jaksoit lukea tänne saakka! Alla lähdeluettelo kaikista artikkelisarjan osista. Aiheeseen liittyvä lukeminen ei ihan heti lopu kesken. Jätä palautetta tai kommentteja ja tsemppiä treeneihin!
Lähteet:
Ahtiainen, J., Pakarinen, A., Kraemer, W. &
Häkkinen, K. (2003). Acute
Hormonal and Neuromuscular Responses and Recovery to Forced vs. Maximum
Repetitions Multiple Resistance Exercises. International Journal of Sports
Medicine, 24(6), 410–418. doi: 10.1055/s-2003-41171
Ahtiainen, J. P.,
Pakarinen, A., Alen, M., Kraemer, W. J., & Häkkinen, K. (2005). Short vs. Long Rest Period Between the Sets in
Hypertrophic Resistance Training: Influence on Muscle Strength, Size, and
Hormonal Adaptations in Trained Men. The Journal of Strength and Conditioning
Research, 19(3), 572. doi: 10.1519/15604.1
Ahtiainen JP, Walker S, Peltonen H, Holviala J,
Sillanpää E, Karavirta L, et al. Heterogeneity in resistance
training-induced muscle strength and mass responses in men and women of
different ages. Age (Omaha). 2016;38:1–13.
Ahmadizad, S., Ghorbani, S., Ghasemikaram, M. &
Bahmanzadeh, M. (2014) Effects of short-term nonperiodized, linear periodized
and daily undulating periodized resistance training on plasma adiponectin,
leptin and insulin resistance. Clinical biochemistry, 47: 417-422.
Alegre, L.,
Jimenez, F., Gonzalo-Orden, J., Acero, R. & Aguado, X. (2006). Effects of
dynamic resistance training on fascicle length and isometric strength. Journal
of sports sciences: 24. 501-8. 10.1080/02640410500189322.
Allen, D. G.,
Whitehead, N. P., & Yeung, E. W. (2005). Mechanisms of stretch-induced
muscle damage in normal and dystrophic muscle: role of ionic changes. The
Journal of Physiology, 567(3), 723–735. doi: 10.1113/jphysiol.2005.091694
Alway, S. E.,
Grumbt, W. H., Stray-Gundersen, J., & Gonyea, W. J. (1992). Effects of
resistance training on elbow flexors of highly competitive bodybuilders.
Journal of Applied Physiology, 72(4), 1512–1521. doi:
10.1152/jappl.1992.72.4.1512
Atherton, P. J.,
Babraj, J., Smith, K., Singh, J., Rennie, M. J., & Wackerhage, H. (2005).
Selective activation of AMPK-PGC-1α or PKB-TSC2-mTOR signaling can explain
specific adaptive responses to endurance or resistance training-like electrical
muscle stimulation. The FASEB Journal, 19(7), 786–788. doi:
10.1096/fj.04-2179fje
Baker, D., Wilson, G. &
Carolyn, R. (1994) Periodization: The effect on strength of manipulating volume
and intensity. Journal of strength and conditioning research, 8: 235-242.
Bamman, M. M.,
Petrella, J. K., Kim, J.-S., Mayhew, D. L., & Cross, J. M. (2007). Cluster analysis tests the importance of myogenic gene
expression during myofiber hypertrophy in humans. Journal of Applied
Physiology, 102(6), 2232–2239. doi: 10.1152/japplphysiol.00024.2007
Benziane, B.,
Burton, T. J., Scanlan, B., Galuska, D., Canny, B. J., Chibalin, A. V., …
Stepto, N. K. (2008). Divergent cell signaling after short-term intensified
endurance training in human skeletal muscle. American Journal of
Physiology-Endocrinology and Metabolism, 295(6). doi:
10.1152/ajpendo.90428.2008
Blazevich, A. J.,
Cannavan, D., Coleman, D. R., & Horne, S. (2007). Influence of concentric
and eccentric resistance training on architectural adaptation in human
quadriceps muscles. Journal of Applied Physiology, 103(5), 1565–1575. doi:
10.1152/japplphysiol.00578.2007
Bondesen, B. A.,
Mills, S. T., & Pavlath, G. K. (2006). The COX-2 pathway regulates growth
of atrophied muscle via multiple mechanisms. American Journal of
Physiology-Cell Physiology, 290(6). doi: 10.1152/ajpcell.00518.2005
Bondesen, B. A.,
Mills, S. T., Kegley, K. M., & Pavlath, G. K. (2004). The COX-2 pathway is
essential during early stages of skeletal muscle regeneration. American Journal
of Physiology-Cell Physiology, 287(2). doi: 10.1152/ajpcell.00088.2004
Brentano, M. &
Martins Kruel, L. (2011) A review on strength exercise-induced muscle damage:
applications, adaption mechanisms and limitations. J Sports Med Phys Fitness,
51(1): 1-10.
Burd, N. A.,
Holwerda, A. M., Selby, K. C., West, D. W. D., Staples, A. W., Cain, N. E., …
Phillips, S. M. (2010). Resistance exercise volume affects myofibrillar protein
synthesis and anabolic signalling molecule phosphorylation in young men. The
Journal of Physiology, 588(16), 3119–3130. doi: 10.1113/jphysiol.2010.192856
Burd, N. A.,
Mitchell, C. J., Churchward-Venne, T. A., & Phillips, S. M. (2012). Bigger
weights may not beget bigger muscles: evidence from acute muscle protein
synthetic responses after resistance exercise. Applied Physiology, Nutrition,
and Metabolism, 37(3), 551–554. doi: 10.1139/h2012-022
Choi J., Takahashi H., Itai Y. (1998) The
difference between effects of ‘power-up type’ and ‘bulk-up type’ strength
training exercises: With special reference to muscle cross-sectional area. Jpn J Phys Fitness Sports Med 47(1), 119-129.
Clarkson, P. M.,
Byrnes, W. C., Mccormick, K. M., Turcotte, L. P., & White, J. S. (1985).
Muscle Soreness And Serum Creatine Kinase Activity Following Isometric,
Eccentric And Concentric Exercise. Medicine & Science in Sports &
Exercise, 17(2), 277. doi: 10.1249/00005768-198504000-00424
Coffey, V. G.,
Zhong, Z., Shield, A., Canny, B. J., Chibalin, A. V., Zierath, J. R., &
Hawley, J. A. (2006). Early signaling responses to divergent exercise stimuli
in skeletal muscle from well-trained humans. The FASEB Journal, 20(1), 190–192.
doi: 10.1096/fj.05-4809fje
Cornelison, D.
& Wold, B. (1997) Single-cell analysis of regulatory gene expression in
quiescent and activated mouse skeletal muscle satellite cells. Advances in
developmental biology, 191(2): 270–283.
Conboy, I. M., Conboy, M. J.,
Wagers, A. J., Girma, E. R., Weissman, I. L., & Rando, T. A. (2005).
Rejuvenation of aged progenitor cells by exposure to a young systemic
environment. Nature, 433(7027): 760–764. doi: 10.1038/nature03260
Dangott, B., Schultz, E.,
& Mozdziak, P. E. (2000). Dietary Creatine Monohydrate Supplementation
Increases Satellite Cell Mitotic Activity During Compensatory Hypertrophy.
International Journal of Sports Medicine, 21(1), 13–16. doi:
10.1055/s-2000-8848
Davitt, P. M., Pellegrino, J.
K., Schanzer, J. R., Tjionas, H., & Arent, S. M. (2014). The Effects of a
Combined Resistance Training and Endurance Exercise Program in Inactive College
Female Subjects. Journal of Strength and Conditioning Research, 28(7),
1937–1945. doi: 10.1519/jsc.0000000000000355
Devaney, J. M., Tosi, L. L.,
Fritz, D. T., Gordish-Dressman, H. A., Jiang, S., Orkunoglu-Suer, F. E., …
Rogers, M. B. (2009). Differences in fat and muscle mass associated with a
functional human polymorphism in a post-transcriptionalBMP2gene regulatory
element. Journal of Cellular Biochemistry, 107(6), 1073–1082. doi:
10.1002/jcb.22209
Dhawan, J., & Rando, T. A.
(2005). Stem cells in postnatal myogenesis: molecular mechanisms of satellite
cell quiescence, activation and replenishment. Trends in Cell Biology, 15(12),
666–673. doi: 10.1016/j.tcb.2005.10.007
Egner, I. M., Bruusgaard, J.
C., Eftestøl, E., & Gundersen, K. (2013). A cellular memory mechanism aids
overload hypertrophy in muscle long after an episodic exposure to anabolic
steroids. The Journal of Physiology, 591(24): 6221–6230. doi:
10.1113/jphysiol.2013.264457
Evans, W. J., & Cannon, J.
G. (1991). The Metabolic Effects of Exercise-Induced Muscle Damage. Exercise
and Sport Sciences Reviews, 19(1). doi: 10.1249/00003677-199101000-00003
Finkenzeller, G., Newsome, W.,
Lang, F., & Häussinger, D. (1994). Increase of c-jun mRNA upon hypo-osmotic
cell swelling of rat hepatoma cells. FEBS Letters, 340(3), 163–166. doi:
10.1016/0014-5793(94)80129-0
Folland, J. &
Williams, A. (2007) The adaptations to strength training - morphological and
neurological contributions to increased strength. Sports medicine, 37 (2):
145-168.
Fonseca, R. M.,
Roschel, H., Tricoli, V., Souza, E. O. D., Wilson, J. M., Laurentino, G. C., …
Ugrinowitsch, C. (2014). Changes in Exercises Are More Effective Than in
Loading Schemes to Improve Muscle Strength. Journal of Strength and
Conditioning Research, 28(11), 3085–3092. doi: 10.1519/jsc.0000000000000539
Fry, C. S., Glynn,
E. L., Drummond, M. J., Timmerman, K. L., Fujita, S., Abe, T., … Rasmussen, B.
B. (2010). Blood flow restriction exercise stimulates mTORC1 signaling and
muscle protein synthesis in older men. Journal of Applied Physiology, 108(5),
1199–1209. doi: 10.1152/japplphysiol.01266.2009
Gardiner NE. Athletics of the ancient world. Oxford:
University Press; 1930.
Glass, D. J.
(2010). PI3 Kinase Regulation of Skeletal Muscle Hypertrophy and Atrophy.
Current Topics in Microbiology and Immunology Phosphoinositide 3-Kinase in
Health and Disease, 267–278. doi: 10.1007/82_2010_78
Goodman, C. A.,
Mayhew, D. L., & Hornberger, T. A. (2011). Recent progress toward
understanding the molecular mechanisms that regulate skeletal muscle mass.
Cellular Signalling, 23(12), 1896–1906. doi: 10.1016/j.cellsig.2011.07.013
Gordon, S. E.,
Kraemer, W. J., Vos, N. H., Lynch, J. M., & Knuttgen, H. G. (1994). Effect
of acid-base balance on the growth hormone response to acute high-intensity
cycle exercise. Journal of Applied Physiology, 76(2), 821–829. doi:
10.1152/jappl.1994.76.2.821
Goto, K., Ishii,
N., Kizuka, T. & Takamatsu, K. (2005) The impact of metabolic stress on
hormonal responses and muscular adaptations. Medicine and science in sports and
exercise, 37(6): 955-963.
Guyton. A. C., Hall J. E. 2011. Textbook of Medical
Physiology. 12th edition. Philadelphia, USA : W.B. Saunders Company
Harries, S. K.,
Lubans, D. R., & Callister, R. (2016). Comparison of resistance training
progression models on maximal strength in sub-elite adolescent rugby union
players. Journal of Science and Medicine in Sport, 19(2), 163–169. doi:
10.1016/j.jsams.2015.01.007
Hackett, D. A.,
Johnson, N. A., & Chow, C.-M. (2013). Training Practices and Ergogenic Aids
Used by Male Bodybuilders. Journal of Strength and Conditioning Research,
27(6), 1609–1617. doi: 10.1519/jsc.0b013e318271272a
Handayaningsih,
A.-E., Iguchi, G., Fukuoka, H., Nishizawa, H., Takahashi, M., Yamamoto, M., …
Takahashi, Y. (2011). Reactive Oxygen Species Play an Essential Role in IGF-I
Signaling and IGF-I-Induced Myocyte Hypertrophy in C2C12 Myocytes.
Endocrinology, 152(3), 912–921. doi: 10.1210/en.2010-0981
Harber, M. P.,
Konopka, A. R., Douglass, M. D., Minchev, K., Kaminsky, L. A., Trappe, T. A.,
& Trappe, S. (2009). Aerobic exercise training improves whole muscle and
single myofiber size and function in older women. American Journal of
Physiology-Regulatory, Integrative and Comparative Physiology, 297(5). doi:
10.1152/ajpregu.00354.2009
Harber, M. P.,
Konopka, A. R., Undem, M. K., Hinkley, J. M., Minchev, K., Kaminsky, L. A., …
Trappe, S. (2012). Aerobic exercise training induces skeletal muscle
hypertrophy and age-dependent adaptations in myofiber function in young and
older men. Journal of Applied Physiology, 113(9), 1495–1504. doi:
10.1152/japplphysiol.00786.2012
Harridge, S.
(2007) Plasticity of human skeletal muscle: Gene expression to in vivo
function. Exp Physiolo. 92: 738-797.
Helms, E., Aragon,
A. & Fitschen, P. (2014). Evidence-based recommendations for natural
bodybuilding contest preparation: Nutrition and supplementation. Journal of the
International Society of Sports Nutrition. 11. 20. 10.1186/1550-2783-11-20.
Hill, M., Wernig,
A., & Goldspink, G. (2003). Muscle satellite (stem) cell activation during
local tissue injury and repair. Journal of Anatomy, 203(1), 89–99. doi:
10.1046/j.1469-7580.2003.00195.x
Hornberger, T. A.,
Chu, W. K., Mak, Y. W., Hsiung, J. W., Huang, S. A., & Chien, S. (2006).
The role of phospholipase D and phosphatidic acid in the mechanical activation
of mTOR signaling in skeletal muscle. Proceedings of the National Academy of Sciences,
103(12), 4741–4746. doi: 10.1073/pnas.0600678103
Izquierdo, M.,
Ibañez, J., González-Badillo, J. J., Häkkinen, K., Ratamess, N. A., Kraemer, W.
J., French, D., Eslava, J., Altadill, A., Asiain X. & Gorostiaga, E. M.
(2006). Differential effects of strength training leading to failure versus not
to failure on hormonal responses, strength, and muscle power gains. Journal of
Applied Physiology, 100(5), 1647–1656. doi: 10.1152/japplphysiol.01400.2005
Jacinto, E., &
Hall, M. N. (2003). TOR signalling in bugs, brain and brawn. Nature Reviews
Molecular Cell Biology, 4(2), 117–126. doi: 10.1038/nrm1018
Kadi, F.,
Eriksson, A., Holmner, S., Butler-Browne, G. & Thornell, L. (1999) Cellular
adaptation of the trapezius muscle in strength-trained athletes. Histochemistry
and cell biology, 111(3): 189-95.
Kadi, F. &
Thornell, L.E. (2000) Concomitant increases in myonuclear and satellite cell
content of female trapezius muscle following strength training. Histochemistry
and cell biology 113, 99-103.
Keogh, J. W.,
Wilson, G. J., & Weatherby, R. E. (1999). A Cross-Sectional Comparison of
Different Resistance Training Techniques in the Bench Press. Journal of
Strength and Conditioning Research, 13(3), 247–258. doi:
10.1519/00124278-199908000-00012
Kohn, T. A., Essén-Gustavsson,
B., & Myburgh, K. H. (2010). Specific muscle adaptations in type II fibers
after high-intensity interval training of well-trained runners. Scandinavian
Journal of Medicine & Science in Sports, 21(6), 765–772. doi:
10.1111/j.1600-0838.2010.01136.x
Kosek, D. J., Kim,
J.-S., Petrella, J. K., Cross, J. M., & Bamman, M. M. (2006). Efficacy of 3
days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in
young vs. older adults. Journal of Applied Physiology, 101(2), 531–544. doi:
10.1152/japplphysiol.01474.2005
Krieger, J. W.
(2010). Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy:
A Meta-Analysis. Journal of Strength and Conditioning Research, 24(4),
1150–1159. doi: 10.1519/jsc.0b013e3181d4d436
Lang, F (2007).
Mechanisms and Significance of Cell Volume Regulation. J Am Coll Nutr., 26:
613s-623s.
Lang, F., Busch,
G. L., Ritter, M., Völkl, H., Waldegger, S., Gulbins, E., & Häussinger, D.
(1998). Functional Significance of Cell Volume Regulatory Mechanisms.
Physiological Reviews, 78(1), 247–306. doi: 10.1152/physrev.1998.78.1.247
Loenneke, J. P.,
Wilson, J. M., Marín, P. J., Zourdos, M. C., & Bemben, M. G. (2011). Low
intensity blood flow restriction training: a meta-analysis. European Journal of
Applied Physiology, 112(5), 1849–1859. doi: 10.1007/s00421-011-2167-x
Loenneke, J. P.,
Thiebaud, R. S., & Abe, T. (2014). Does blood flow restriction result in
skeletal muscle damage? A critical review of available evidence. Scandinavian
Journal of Medicine & Science in Sports, 24(6). doi: 10.1111/sms.12210
Lynn, R. &
Morgan, D. (1994) Decline running produces more sarcomeres in rat vastus
intermedius muscle fibers than does incline running. Journal of applied
physiology: 77: 1439-1444.
Malm, C. (2001).
Exercise-induced muscle damage and inflammation: fact or fiction? Acta
Physiologica Scandinavica, 171(3), 233–239. doi:
10.1046/j.1365-201x.2001.00825.x
Macdougall, J. D.,
Gibala, M., Tarnopolsky, M., Macdonald, J., Interisano, S., & Yarasheski,
K. (1995). The Time Course For Elevated Muscle Protein Synthesis Following
Heavy Resistance Exercise. Medicine & Science in Sports & Exercise,
27(Supplement). doi: 10.1249/00005768-199505001-00367
MacDougall, D. Ward, G., Sale, D. & Sutton, J.
(1977) Biochemical adaptation of human skeletal muscle to heavy resistance
training and immobilization. Journal of applied physiology:
respiratory, environmental and exercise physiology: 43(4): 700-3.
Macdougall, J. D., Sale, D. G., Alway, S. E., &
Sutton, J. R. (1984). Muscle fiber number in biceps brachii in bodybuilders and
control subjects. Journal of Applied Physiology, 57(5), 1399–1403. doi:
10.1152/jappl.1984.57.5.1399
Mascher, H., Andersson, H., Nilsson, P.-A., Ekblom,
B., & Blomstrand, E. (2007). Changes in signalling pathways regulating
protein synthesis in human muscle in the recovery period after endurance
exercise. Acta Physiologica, 191(1), 67–75. doi: 10.1111/j.1748-1716.2007.01712.x
Masuda K., Choi J.Y., Shimojo H., Katsuta S. (1999) Maintenance of myoglobin concentration in human skeletal muscle
after heavy resistance training. European
Journal of Applied Physiology and Occupational Physiology 79(4), 347-352. [PubMed] [Google Scholar]
Martineau, L. & Gardiner,
P. (2002) Skeletal muscle is sensitive to the tension-time integral but not to
the rate of change of tension, as assessed by mechanically induced signaling.
Journal of biomechanic, 35: 657-663.
Mchugh, M. P. (2003). Recent advances in the
understanding of the repeated bout effect: the protective effect against muscle
damage from a single bout of eccentric exercise. Scandinavian Journal of
Medicine and Science in Sports, 13(2), 88–97. doi:
10.1034/j.1600-0838.2003.02477.x
Mcpherron, A. C., Lawler, A.
M., & Lee, S.-J. (1997). Regulation of skeletal muscle mass in mice by a
new TGF-p superfamily member. Nature, 387(6628), 83–90. doi:
10.1038/387083a0
Mikkola,
J., Rusko, H., Izquierdo, M., Gorostiaga, E., & Häkkinen, K. (2012). Neuromuscular and Cardiovascular Adaptations During
Concurrent Strength and Endurance Training in Untrained Men. International
Journal of Sports Medicine, 33(09), 702–710. doi: 10.1055/s-0031-1295475
Moore, D., Phillips, S., Babraj, J., Smith, K. & Rennie, M. (2005) Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. American journal of physiology - endocrinology andmetabolism, 288: E1153–E1159.
Monteiro, A., Aoki., M.,
Evangelista,A., Alveno, D., Monteiro, G., Picarro Ida, C. & Ugrinowitcch,
C. (2009) Nonlinear periodization maximizes strength gains in split resistance
training routines. Journal of strength and conditioning research, 23: 1321-1326.
Netreba, A., Popov, D., Bravyy,
Y., Lyubaeva, E., Terada, M., Ohira, T.,
Okabe, H., Vinogradova, O.& Ohira, Y. (2013). Responses of knee extensor
muscles to leg press training of various types in human. Rossiĭskii
fiziologicheskiĭ zhurnal imeni I.M. Sechenova / Rossiĭskaia akademiia nauk. 99.
406-16.
Nguyen, H. X., & Tidball,
J. G. (2003). Null Mutation of gp91phoxReduces Muscle Membrane Lysis During
Muscle Inflammation in Mice. The Journal of Physiology, 553(3), 833–841. doi:
10.1113/jphysiol.2003.051912
Nosaka, K., Lavender, A.,
Newton, M. & Sacco, P. (2003) Muscle damage in resistance training – is muscle
damage necessary for strength gain and muscle hypertrophy? International
journal of sport and health science, 1(1):1-8.
Nuckols, G. (2018)
Periodization: What the data say. https://www.strongerbyscience.com/periodization-data/
Ogasawara, R., Kobayashi, K.,
Tsutaki, A., Lee, K., Abe, T., Fujita, S., … Ishii, N. (2013). mTOR signaling
response to resistance exercise is altered by chronic resistance training and
detraining in skeletal muscle. Journal of Applied Physiology, 114(7), 934–940.
doi: 10.1152/japplphysiol.01161.2012
Pescatello, L. S., Devaney, J.
M., Hubal, M. J., Thompson, P. D., & Hoffman, E. P. (2013). Highlights from
the Functional Single Nucleotide Polymorphisms Associated with Human Muscle
Size and Strength or FAMuSS Study. BioMed Research International, 2013, 1–11.
doi: 10.1155/2013/643575
Petrella, J. K., Kim, J.-S.,
Mayhew, D. L., Cross, J. M., & Bamman, M. M. (2008). Potent myofiber
hypertrophy during resistance training in humans is associated with satellite
cell-mediated myonuclear addition: a cluster analysis. Journal of Applied
Physiology, 104(6), 1736–1742. doi: 10.1152/japplphysiol.01215.2007
Pinto, R., Gomes, N.,
Radaelli, R., Botton, C., Brown, L. & Bottaro, M. (2011). Effect of Range
of Motion on Muscle Strength and Thickness. Journal of strength and
conditioning research / National Strength & Conditioning Association. 26.
2140-5. 10.1519/JSC.0b013e31823a3b15.
Poliquin, C. (1988) Five steps
to increasing the effectiveness of your strength training program. Journal of
strength and conditioning association. 10: 34-39.
Prestes, J., Frollini, A. B.,
Lima, C. D., Donatto, F. F., Foschini, D., Marqueti, R. D. C., … Fleck, S. J.
(2009). Comparison Between Linear and Daily Undulating Periodized Resistance
Training to Increase Strength. Journal of Strength and Conditioning Research,
23(9), 2437–2442. doi: 10.1519/jsc.0b013e3181c03548
Qaisar, R., Bhaskaran, S.
& Remmen, H. (2016). Muscle
fiber type diversification during exercise and regeneration. Free Radical
Biology and Medicine. 98. 10.1016/j.freeradbiomed.2016.03.025.
Radaelli, R. J Fleck, S.,
Leite, T., Leite, R., Pinto, R., Fernandesm L. Simão, R. (2014). Dose Response
of 1, 3 and 5 Sets of Resistance Exercise on Strength, Local Muscular Endurance
and Hypertrophy. The Journal of Strength and Conditioning Research. 29.
10.1519/JSC.0000000000000758.
Ratamess, N. A., Falvo, M. J.,
Mangine, G. T., Hoffman, J. R., Faigenbaum, A. D., & Kang, J. (2007). The
effect of rest interval length on metabolic responses to the bench press
exercise. European Journal of Applied Physiology, 100(1), 1–17. doi:
10.1007/s00421-007-0394-y
Schoenfeld, B. (2010) The
mechanism of muscle hypertrophy and their application to resistance training.
Journal of strength and conditioning research, 24: 2857-2872.
Schoenfeld, BJ. Science and Development of Muscle Hypertrophy. Champaign, IL; Human Kinetics, 2016.
Schoenfeld, B, Ogborn, D.
& Krieger, J. (2017) Dose-response relationship between weekly resistance
training volume and increases in muscle mass: A systematic review and
meta-analysis, Journal of Sports Sciences, 35:11, 1073-1082, DOI:
10.1080/02640414.2016.1210197
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., Sonmez, G. T., & Alvar, B. A.
(2014). Effects of Different Volume-Equated Resistance Training Loading
Strategies on Muscular Adaptations in Well-Trained Men. Journal of Strength and
Conditioning Research, 28(10), 2909–2918. doi: 10.1519/jsc.0000000000000480
Schoenfeld, B, Grgic, J.,
Ogbord, D. & Krieger, J. (2017) Strength and hypertrophy adaptations
between low- versus high-load resistance training: A systematic review and
meta-analysis. The Journal of strength and conditioning research, Ahead of
Print DOI: 10.1519/JSC.0000000000002200
Schoenfeld, B., Contreras, B.,
Vigotsky, A., Ogborn, D., Fontana, F. & Tiryaki-Sonmez, R. (2016). Upper
body muscle activation during low-versus high-load resistance exercise in the
bench press. 24. 217-224. 10.3233/IES-160620.
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., & Tiryaki-Sonmez, G. (2015).
Influence of Resistance Training Frequency on Muscular Adaptations in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(7),
1821–1829. doi: 10.1519/jsc.0000000000000970
Schoenfeld, B. J., Peterson,
M. D., Ogborn, D., Contreras, B., & Sonmez, G. T. (2015). Effects of Low-
vs. High-Load Resistance Training on Muscle Strength and Hypertrophy in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(10), 2954–2963.
doi: 10.1519/jsc.0000000000000958
Schoenfeld, B. J., Ogborn, D.
I., & Krieger, J. W. (2015). Effect of Repetition Duration During
Resistance Training on Muscle Hypertrophy: A Systematic Review and
Meta-Analysis. Sports Medicine, 45(4), 577–585. doi: 10.1007/s40279-015-0304-0
Schoenfeld, B. J., Contreras,
B., Krieger, J., Grgic, J., Delcastillo, K., Belliard, R., & Alto, A.
(2019). Resistance Training Volume Enhances Muscle Hypertrophy but Not Strength
in Trained Men. Medicine & Science in Sports & Exercise, 51(1), 94–103.
doi: 10.1249/mss.0000000000001764
Schoenfeld, B.J. & Grgic,
J. (2020) Effects of range of motion on muscle development during resistance
training interventions: A systematic review. SAGE open medicine, 8: 1-8. https://doi.org/10.1177/2050312120901559
Schuenke, M. D., Herman, J.
R., Gliders, R. M., Hagerman, F. C., Hikida, R. S., Rana, S. R., … Staron, R.
S. (2012). Early-phase muscular adaptations in response to slow-speed versus
traditional resistance-training regimens. European Journal of Applied
Physiology, 112(10), 3585–3595. doi: 10.1007/s00421-012-2339-3
Selye, H. (1950). Stress and
the General Adaptation Syndrome. Bmj, 1(4667), 1383–1392. doi:
10.1136/bmj.1.4667.1383
Serrano, A. L., Baeza-Raja,
B., Perdiguero, E., Jardí, M., & Muñoz-Cánoves, P. (2008). Interleukin-6 Is
an Essential Regulator of Satellite Cell-Mediated Skeletal Muscle Hypertrophy.
Cell Metabolism, 7(1), 33–44. doi: 10.1016/j.cmet.2007.11.011
Simão, R., Spineti, J.,
Salles, B. F. D., Matta, T., Fernandes, L., Fleck, S. J., … Strom-Olsen, H. E.
(2012). Comparison Between Nonlinear and Linear Periodized Resistance Training.
Journal of Strength and Conditioning Research, 26(5), 1389–1395. doi:
10.1519/jsc.0b013e318231a659
Snyder, B. J., & Leech, J.
R. (2009). Voluntary Increase in Latissimus Dorsi Muscle Activity During the
Lat Pull-Down Following Expert Instruction. Journal of Strength and
Conditioning Research, 23(8), 2204–2209. doi: 10.1519/jsc.0b013e3181bb7213
Snyder, B. J., & Fry, W.
R. (2012). Effect of Verbal Instruction on Muscle Activity During the Bench
Press Exercise. Journal of Strength and Conditioning Research, 26(9), 2394–2400.
doi: 10.1519/jsc.0b013e31823f8d11
Stewart, C. & Rittweger,
J. (2006) Adaptive processes in skeletal muscle: molecular regulators and
genetic influences. Journal of musculoskeletal and neuronal interactions: 6(1): 73-86.
Stone, M., O’Bryant, H. &
Garhammer J. (1981) Hypothetical model for strength training. Journal of sports
medicine and physical fitness, 21: 342-351.
Stone, M. H., Potteiger, J.
A., Pierce, K. C., Proulx, C. M., Obryant, H. S., Johnson, R. L., & Stone,
M. E. (2000). Comparison of the Effects of Three Different Weight-Training
Programs on the One Repetition Maximum Squat. The Journal of Strength and
Conditioning Research, 14(3), 332. doi:
10.1519/1533-4287(2000)014<0332:coteot>2.0.co;2
Takarada,
Y., Takazawa, H., Sato, Y., Takebayashi, S., Tanaka, Y., & Ishii, N.
(2000). Effects of
resistance exercise combined with moderate vascular occlusion on muscular
function in humans. Journal of Applied Physiology, 88(6), 2097–2106. doi:
10.1152/jappl.2000.88.6.2097
Takarada, Y., Nakamura, Y.,
Aruga, S., Onda, T., Miyazaki, S., & Ishii, N. (2000). Rapid increase in plasma growth hormone after
low-intensity resistance exercise with vascular occlusion. Journal of Applied
Physiology, 88(1), 61–65. doi: 10.1152/jappl.2000.88.1.61
Tee, J. C., Bosch, A. N.,
& Lambert, M. I. (2007). Metabolic Consequences of Exercise-Induced Muscle
Damage. Sports Medicine, 37(10), 827–836. doi: 10.2165/00007256-200737100-00001
Terzis, G., Spengos, K., Mascher,
H., Georgiadis, G., Manta, P. &
Blomstrand, E. (2010). The degree of p70S6k and S6 phosphorylation in human
skeletal muscle in response to resistance exercise depends on the training
volume. European journal of applied physiology. 110. 835-43.
10.1007/s00421-010-1527-2.
Thomas, G., & Hall, M. N.
(1997). TOR signalling and control of cell growth. Current Opinion in Cell
Biology, 9(6), 782–787. doi: 10.1016/s0955-0674(97)80078-6
Uchiyama, S., Tsukamoto, H.,
Yoshimura, S., & Tamaki, T. (2006). Relationship between oxidative stress in muscle tissue
and weight-lifting-induced muscle damage. Pflügers Archiv - European Journal of
Physiology, 452(1), 109–116. doi: 10.1007/s00424-005-0012-y
Wernbom, M., Augustsson, J.
& Thomee, R. (2007). The Influence of Frequency, Intensity, Volume and Mode
of Strength Training on Whole Muscle Cross-Sectional Area in Humans. Sports
medicine (Auckland, N.Z.). 37. 225-64. 10.2165/00007256-200737030-00004.
Willoughby, D. (1993) The
effects of mesocycle-length weight training programs involving periodization an
partially equated volumes on upper and lower body strength. Journal of strength
and conditioning research, 7: 2-8.
Wilson, J. M., Lowery, R. P.,
Joy, J. M., Loenneke, J. P., & Naimo, M. A. (2013). Practical Blood Flow
Restriction Training Increases Acute Determinants of Hypertrophy Without
Increasing Indices of Muscle Damage. Journal of Strength and Conditioning
Research, 27(11), 3068–3075. doi: 10.1519/jsc.0b013e31828a1ffa
Wilson, J. M., Marin, P. J.,
Rhea, M. R., Wilson, S. M., Loenneke, J. P., & Anderson, J. C. (2012).
Concurrent Training. Journal of Strength and Conditioning Research, 26(8),
2293–2307. doi: 10.1519/jsc.0b013e31823a3e2d
Wulf, G. (2013). Attentional
focus and motor learning: a review of 15 years. International Review of Sport
and Exercise Psychology, 6(1), 77–104. doi: 10.1080/1750984x.2012.723728
Yang, S. Y., & Goldspink,
G. (2002). Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in
myoblast proliferation and differentiation. FEBS Letters, 522(1-3), 156–160.
doi: 10.1016/s0014-5793(02)02918-6
Zanou, N., & Gailly, P.
(2013). Skeletal muscle hypertrophy and regeneration: interplay between the
myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs)
pathways. Cellular and Molecular Life Sciences, 70(21), 4117–4130. doi:
10.1007/s00018-013-1330-4
Zanchi, N. E., & Lancha,
A. H. (2007). Mechanical stimuli of skeletal muscle: implications on
mTOR/p70s6k and protein synthesis. European Journal of Applied Physiology,
102(3), 253–263. doi: 10.1007/s00421-007-0588-3
Maksimivoima toimii pohjana monelle muulle ominaisuudelle. Voimaa tarvitaan muun muassa siirtämään urheilijan omaa kehoa, vastustajaa tai välinettä. Voimaa tarvitaan erityisesti kuulantyönnössä, jossa pitää siirtää pienessä betoniympyrässä rautapallo mahdollisimman kauas itsestä. Knoppitietona miesten kuulantyönnön ME-työntö 23,12 m on kuulaan tuotettuna voimana noin hevosvoiman luokkaa. Yksi hevosvoima vastaa suunnilleen 735,5 wattia. Kokonaisteho on luontaisesti tätä paljon suurempi, jopa kahdeksankertainen. (Yrjölä, 2000.)
Muissa
lajeissa tarvittavan voiman määrä vaihtelee lajikohtaisesti. Tähän vaikuttaa
paljolti, missä asennoissa voimaa tuotetaan, kuinka nopeasti sitä tulee
tuottaa, millaisissa liikkeissä sitä tulee tuottaa ja kuinka monta kertaa
peräkkäin. (Häkkinen & Ahtiainen, 2016.)
Voima voidaan jakaa nopeusvoimaan, maksimivoimaan ja kestovoimaan. Maksimivoimalla tarkoitetaan suurinta voimatasoa, jonka lihas tai lihasryhmä kykenee tuottamaan tahdonalaisessa kertasupistuksessa. Tässä tekstissä pureudutaan puhtaasti maksimivoimaharjoitteluun. Tiivistetysti maksimivoimaharjoittelun tarkoitus on kehittää hermolihasjärjestelmän kykyä tuottaa voimaa ja tukikudosten rasituskestävyyttä. (Häkkinen & Ahtiainen, 2016.)
Kuulantyöntö ja
maksimivoimaharjoittelu
Kuulantyöntö on loistava esimerkki maksimivoiman merkityksestä lajissa. Kuulantyöntäjä tarvitsee todella kovat maksimivoimatasot, jotta naisilla 4-kiloinen ja miehillä 7,26-kiloinen kuula liikahtaa ringistä mahdollisimman pitkälle. Voimantuottoaika kuulantyönnössä on vain noin 1,3 - 1,5 sekuntia, ja itse työntövaihe kestää siitä 0,2 - 0,25 sekuntia (Yrjölä, 2000). Voima pitää siis kyetä myös tuottamaan nopeasti. Suuresta määrästä voimaa ei ole hyötyä, jos sitä ei pysty tuottamaan räjähtävästi rautapalloon. Kuulantyöntökilpailun aikana voimaa tuotetaan vain kuusi kertaa, ja jokainen suoritus on asyklinen eli tapahtuu vain kerran. Kuulantyöntö on siksi loistava tarkastelunkohde, koska maksimivoimatasot tulee olla äärimmäisen korkealla, ja voima tulee tuottaa nopeasti.
Verrattuna vaikkapa voimanostajiin kuulantyöntäjän tulee olla paljon räjähtävämpi ja monipuolisempi urheilija. Voimannostajan ei tarvitse nostaa painoa nopeasti, kunhan paino vain nousee. Kuulantyöntäjän taas pitää lyhyessä ajassa tuottaa voimaan kuulaa, mutta toisaalta kuulantyöntäjien ei tarvitse periaattessa olla maksimivoimatasoiltaan yhtä voimakkaita kuin voimannostajien. Kuulantyönnön voimantuottoaika on hyvin lyhyt. Se on vain 0,2 – 0,25 sekuntia kun tahdonalaisen maksimaalisen voimatason saavuttaminen vie jopa 0,5 – 2,5 sekuntia riippuen lihaksesta, liikkeestä, harjoitustaustasta ja geeneistä (Mero, Ahtiainen & Häkkinen, 2007). Tämän takia voimanostajien harjoittelu on hieman erilaista. Painonnostoharjoittelu näyttäisi sopivan erityisen hyvin kuulantyöntäjille. Se johtuu painonnostoliikkeiden räjähtävämmästä luonteesta verrattuna voimanostoon.
No kuinka vahva
kuulantyöntäjän tulee sitten olla?
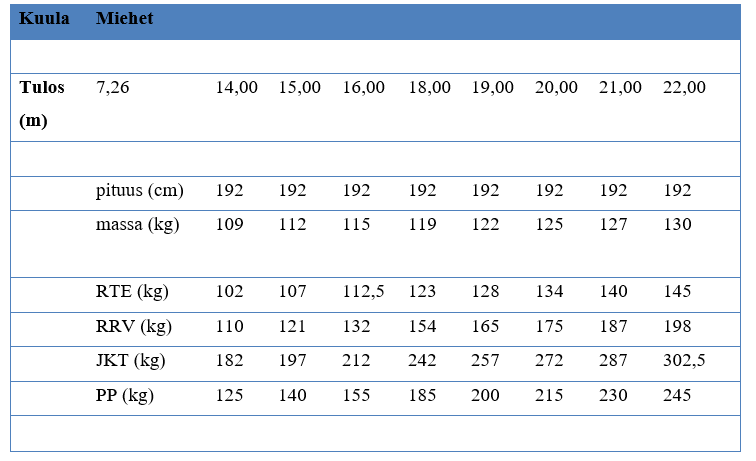
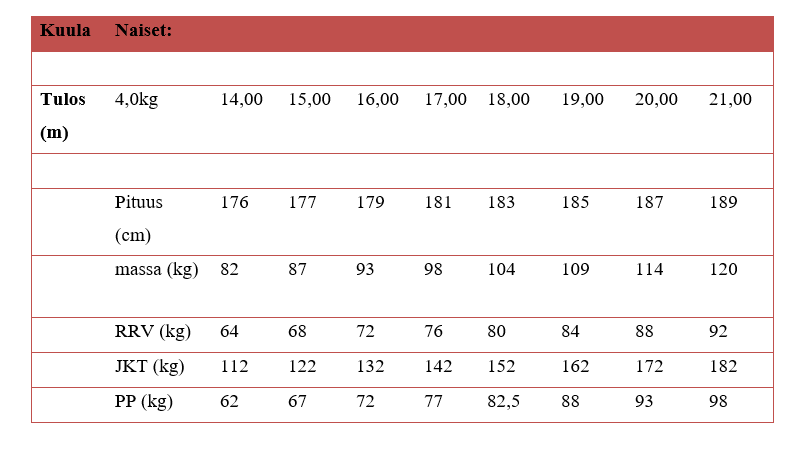
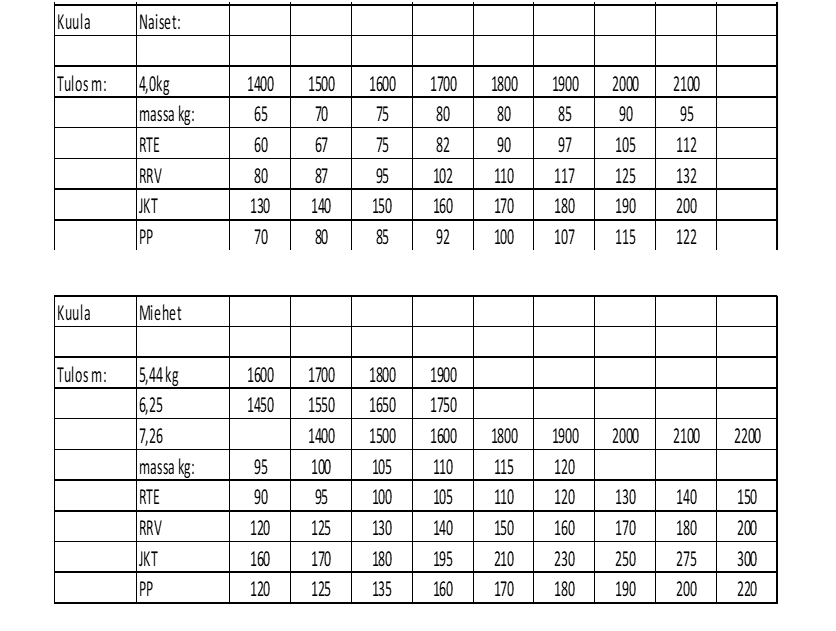
Tarvittavia
voimatasoja on yrittänyt lisäkseni määritellä Auvinen (2011) (Räntilä, 2015)
(taulukot 1 ja 2.). Ylhäältä voi lukea halutun tuloksen ja siihen vaadittavat
voimatasot. Taulukko on suuntaa antava, ja sitä voi käyttää apuna urheilijan
harjoittelun ohjelmoinnissa. Urheilijan yksilölliset vahvuudet ja heikkoudet
kannattaa huomioida tarkkaan.
Kaikki
ominaisuudet heittolajeissa ovat rakennettavissa, ja melkein jokaisesta
innostuneesta heittäjästä olisi mahdollisuus saada hyvän tason heittäjä
Suomessa. Ominaisuuksien tasainen rakentaminen on kuitenkin valmennuksellisesti
vaikeaa, ja se vaatii pitkäjänteistä työtä vuodesta toiseen. Fyysiset
ominaisuudet eivät kehity hetkessä tai nopeasti vaan tasaisesti ja hitaasti.
Ominaisuusnormistot (taulukot 1 ja 2) ovat oiva väline nuorelle heittäjälle
seuraamaan omaa kehitystään. Omien vahvuuksien ja heikkouksien löytäminen
helpottuu taulukoiden avulla. Taulukoita luetaan niin, että tulokset ovat ylärivissä
ja fyysiset ominaisuudet pystyrivissä. Esimerkiksi jos mieskuulantyöntäjä
haluaa työntää yli 20 metriä niin tulisi hänen rinnallevetonsa olla 175 kg ja
penkkipunnerruksen 215 kg.
Karoliina Lundahl, suomalainen painonnoston maailmanmestari vuosilta 1994 ja 1998, edusti Suomea Atlantan olympiakisoissa vuonna 1996 myös kuulantyönnössä. Hänen voimatasonsa olivat erittäin korkeat ja onkin spekuloitu, että hänen voimatasonsa olisivat riittäneet huomattavasti parempaankin kuulantyöntötulokseen. Lundahlin ennätys on 18,28 m. Hänen painonnostoennätyksillään (tempaus 107 kg ja työntö 127 kg) olisi Auvisen taulukon mukaan pitänyt työntää 20 metriä (katso taulukot 1 ja 2).
Nykytyöntäjistä esimerkiksi Suvi Helin on yhdistänyt painonnoston ja kuulantyönnön huipputasolla. Hänen ennätyksensä on 15,96 m. Suvi Helinin ennätykset vuonna 2015 olivat tempauksessa 85 kg ja työnnössä 105 kg. Näillä hänen olisi pitänyt työntää yli 17 metriä, mutta hän on työntänyt “vain” 15,96 m. Miksi nämä naiset eivät ole työntäneet pidemmälle hurjista voimatasoista huolimatta? Selitys löytyy yksinkertaisesti tekniikasta. Voima tulee tuottaa välineeseen lyhyessä ajassa vaativien liikesarjojen kautta. Nyt voimaa ei ole saatu tuotettua välineeseen. Tämän takia kuulantyöntäjä viettää harjoitusajasta karkeasti vain puolet salilla, joissakin tapauksissa jopa huomattavasti vähemmänkin (Zatsiorsky & Kramer, 2006).
Painonnoston ja kuulantyönnön yhdistäminen huipputasolla on siis erittäin haastavaa. Lajit vaativat hyvin erilaista erikoistumista harjoittelun ja kehon jänteiden sekä nivelten adaptaatioiden suhteen. Harjoittelu kuulantyönnön huipputasolla vaatii hyvin paljon työntämistä ja erilaisia drillejä sekä rotaatiovoiman kehittämistä. Painnonostoharjoittelu vaatii kasvavan harjoittelumäärän omalta osaltaan. Näiden yhdistäminen on ajankäytöllisesti ja kehon jaksamisen takia hyvin vaikeaa. Toisaalta nuorille urheilijoille näiden lajien yhdistäminen sopii erinomaisesti.
Taulukkoa 1 varten on kerätty aineistoa suomalaisista huipuista ja sen takia esimerkiksi miesten kuulantyönnössä pituus ei muutu. Suomalaisessa kuulantyönnössä on vaikuttanut paljon pyörähtäjiä ja heidän antropometriansa ei ole yhteydessä kuulantyöntö tuloksen kanssa. Toisin kuin pakittajilla. Jos pakittajia olisi aineistossa enemmän, olisi pituus varmasti merkitsevämmässä roolissa. Lisäksi Suomessa on aika vähän huippunaistyöntäjiä ollut, joten naisten taulukon aineisto on selvästi painottunut 14 metrin ja 17 metrin välille. Uskon itse, että yli 17 metrin työnnöt vaativat hiukan kovemmat maksimivoimatasot kuin mitä taulukossa nyt on.
Kuten kaikkien voimaharjoittelijoiden kuulantyöntäjänkin tulee käyttää 2-3 vuotta tekniikoiden hiomiseen, ennen kuin nousujohteinen voimaharjoittelu voi alkaa. Tätä tekniikkaharjoittelua on helppoa tehdä 11 - 15-vuotiaana, jolloin harjoituksiin voi yhdistää liikkuvuuden, kestovoiman ja nopeusvoiman elementtejä. Vanhemmalla iällä, kun voimaa jo on, tekniikoiden rakentaminen alusta on yleensä varsin mälsää puuhaa. Kuulantyöntäjälle parhaat liikkeet riippuen työntäjän antropometriasta ovat tempausvariaatiot, rinnallevetovariaatiot, työntövariaatiot, kyykkyvariaatiot, maastavetovariaatiot ja erilaiset apuliikkeet. Kuulantyönnössä korostuu jalkojen vertikaalinen voimantuotto ja yläkropan horisontaalinen voimantuotto, jonka kehittämiseen esimerkiksi penkkipunnerrus on hyvä liike.
Harjoittelun yksi tärkeimpiä tekijöitä on nousujohteisuus. Harjoitusmäärien tasainen nostaminen voimaharjoittelussa tulee tehdä tasaisesti monen vuoden aikana. Suuria voimaominaisuuksia ei saavuteta nopeasti tai lyhyen aikavälin runsailla volyymilisäyksillä. Lisäksi äkkinäiset volyymilisäykset altistavat urheilijan turhalle loukkaantumisriskille.
Volyymi saadaan kertomalla toistot
sarjalla ja painolla, esimerkiksi 10 sarjaa 10 toistoa 100 kilolla tekee
yhteensä 10 000 kiloa eli 10 tonnia. Yleensä volyymiin lasketaan vain ne sarjat,
jotka tehdään yli 70 % intensiteetillä lasketusta ykkösmaksimista. Kuulantyöntäjän tulee ohjeellisesti nostaa
minimissään 900 tonnia vuodessa ja maksimissaan 1450 tonnia vuodessa.
Valmistavalla kaudella ohjeellisesti tonnit vaihtelevat 24 - 40 välillä yhdellä
harjoitusviikolla ja kilpailukaudella 8 - 12 tonnin välillä. (Bompa & Haff, 2009)
Harjoittelun kehitystä voi seurata myös laskemalla tehdyt sarjat, jotka ylittävät tietyn tuntemuksen rajan (esim. RPE taulukko) tai sarjat, joissa nopeus tippui tietyn prosenttiosuuden verran (esim. 20 % maksiminopeudesta, tämä vaatii tosin nopeuden mittaamisen).
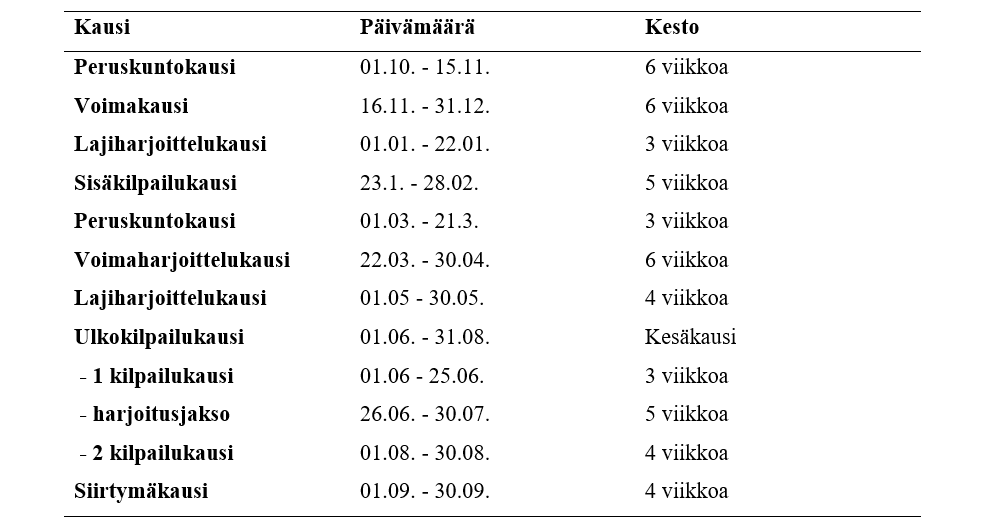
TAULUKKO 3: Kuulantyöntäjän klassinen ja paljon käytetty harjoitusvuoden jaottelu. Vaikka painopisteet on jaoteltu, tulee voimaharjoittelu ja työntäminen pitää mukana kaikilla harjoittelukausilla. (Mukailtu Yrjölä, 2000). Nykyään suositaan myös paljon blokkiperiodisaatiota klassisen periodisaation rinnalla.
Hypertrofisella harjoittelulla tarkoitetaan lihassolun poikkipinta-alan kasvuun tähtäävää harjoittelua. Harjoittelujakson tavoitteena voi olla rasvattoman massan lisääminen, kudoksien vahvistaminen, mahdollisesti myös rasvamassan vähentäminen ja työkapasiteetin nostaminen, Tämä luo pohjat intensiteetin ja volyymin nostolle. Lisäksi luodaan hermolihasjärjestelmään tasapaino ja ehkäistään puolieroja ja vaikutetaan loukkaantumisten ehkäisyyn. (Bompa & Haff, 2009.)
Hypertrofiaa voidaan saavuttaa laajalla skaalalla erilaisia harjoitusohjelmia. Tärkeimmät vaikuttavat muuttujat harjoitusohjelmat lopputulokseen ovat kuorma, intensiteetti, nopeus, volyymi, sarjojen välinen lepo, liiketyyppi ja aktiivisen lihasmassan määrä. Volyymin ajatellaan olevan erittäin tärkeä tekijä morfologisten adaptaatioiden saavuttamisiksi (Krieger, 2010). Volyymin nosto harjoitusohjelmassa on helppo progressiotapa hypertrofisen jakson aikana. Yksi lihaskasvun tärkeimmistä ärsykkeistä on mekaaninen kuormitus (Schiaffino et al., 2013). Myös metabolisella rasituksella ajatellaan olevan rooli hypertrofiassa (Schoenfeld, 2013) ja myös jossain määrin voimassa (Rooney et al., 1994; Schott et al., 1995). Näiden kummankin yhdistäminen voi olla optimaalista anatomisten adaptaatioiden saavuttamiseksi.
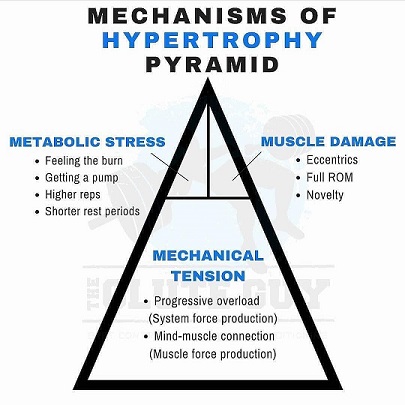
KUVA 1: Bret Contreras kuvasi hyvin mekaanisen ja metabolisen rasituksen sekä lihasvaurioiden suhdetta. Hypertrofisessa harjoittelussa mekaaninen jännitys ja progressiivinen kuormitus ovat tärkeintä. Metabolisella kuormituksella ja lihasvauriolla on osuus kokonaisuudessa, mutta se on pienehkö.
Maksimivoimaharjoittelulla kehitetään sitä maksimaalista voimaa, mikä kyetään tuottamaan. Tällöin yleensä harjoittelu keskittyy hermostollisen ja lihakseen liittyvän kapasiteetin maksimaalisten adaptaatioiden saavuttamiseen. Kehittyäkseen maksimivoima tarvitsee korkean intensiteetin.
Maksimaalisella yrittämisellä (aggressiivinen nopeusvoimaperiaate) rekrytoidaan mahdollisimman täydellisesti nopeat motoriset yksiköt eli nopeat liikehermot ja niiden päissä olevat nopeasti supistuvat lihassolut. Tämä on erittäin tärkeää maksimivoimajaksolla kuin myös sitä seuraavalla nopeusvoimajaksolla.
TAULUKKO 4: Miten voimaharjoittelu voitaisiin käytännössä ohjelmoida.
Lisää
räjähtävää voimaa
Räjähtävävoiman ja pikavoima jaksolla on tarkoitus oppia tuottamaan voimaa mahdollisimman paljon lyhyessä ajassa. Ohjelmoinnista riippuen harjoittelussa voi tehdä myös yhdistettyä voima- ja nopeusharjoittelua. Kuulantyöntäjän harjoittelussa tulee muistaa vielä lajivoiman lisääminen ohjelmointiin. Lajivoiman kehittäminen tapahtuu yleensä eripainoisia välineitä työntämällä. Taulukosta 4 voi katsoa karkean esimerkin vuositason ohjelmoinnista kokeneemmalle urheilijalle.
Räjähtävä voima tarkoittaa kykyä käyttää maksimaalista voimaa mahdollisimman lyhyessä ajassa. Parhailla kuulantyöntäjillä 21 metrin työnnöissä on mitattu heittokäden käsityönnön voimaksi 50-60 kg. Näiden työntäjien maksimaalinen käsien ojennus - eli penkkipunnerrustulos - vaihtelee tyypillisesti 220-240 kilon välillä eli 110-120 kg kättä kohden. Itse lajisuorituksessa on tällöin onnistuttu tuottamaan noin 50 % maksimaalisesta käytössä olevasta voimasta. Heittolajeissa voi parantaa voimantuottoa kahdella tavalla. Ensinnäkin voi lisätä maksimaalista voimaa. Toinen tapa on lisätä voimantuottonopeutta. Heittäjän uran alussa lapsuudessa ja nuoruudessa voi olla viisasta kehittää maksimaalista voimaa ja yleisesti lihaksistoa unohtamatta nopeuden herkkyysvaiheita. Tämä antaa laajan pohjan voimantuottonopeusharjoittelulle. Toisinpäin tehtynä pelkkää nopeutta kehittäen maksimaalista voimaa ei saada enää uralla riittävästi käyttöön. Tehokkainta on näiden kummankin yhdistäminen optimaalisesti ja yksilöllisesti. (Zatsiorsky & Kramer 2006.)
On tärkeää tehdä maksimivoimajakso ennen
räjähtävän voiman jaksoa, koska tehon lisäys voi tapahtua niin maksimaalisen
voiman kuin nopeuden lisäyksenä tai molempien kasvulla. Jakson pituus voi
vaihdella yhdestä kolmeen kuukauteen, riippuen kisataanko hallikaudella.
Räjähtävän voiman/pikavoiman kausi tulee sijoittaa kilpailuun valmistavalle
kaudelle. Sen tarkoituksena on muuntaa saavutetut korkeammat voimatasot lajin
käyttöön. On kuitenkin tärkeä muistaa ylläpitää maksimivoimatasoja, koska
muuten nopeus kärsii. (Bompa & Haff, 2009.)
Siirtymävaihe on kuulantyöntäjälle hyvin tärkeä. Tämä jakso ajoittuu heti kilpailukauden jälkeen ennen uuden kauden aloittamista. Jakson pituus vaihtelee urheilijan tason mukaan kahdesta viikosta jopa kuuteen viikkoon. Tärkein tavoite on fyysisesti ja psyykkisen väsymyksen poistaminen. Siirtymävaiheen aikana tulee olla aktiivinen, pelkkä passiivinen lepo ei välttämättä riitä palautumiseen. Suositeltavaa on myös antaa paljon käytetyille nivelille ja jänteille lepoa. Esimerkiksi työntökädelle ei tehdä siirtymäkaudella työntöjä (penkki, lajisuoritukset, pystypunnerrus ym.). Selän kompressio- ja leikkausvoimia selälle on lisäksi hyvä välttää (kyykky, maastaveto ja vastaavat liikkeet).
Viimeinen ominaisuuksien
nosto: herkistely
Herkistely on kilpailukaudella erittäin tärkeää. Tarkoituksena on poistaa harjoittelun aiheuttama väsymys mutta ylläpitää ja jopa nostaa esiin tiettyjä ominaisuuksia. Taulukosta 5 voi nähdä, että karkeasti harjoittelun volyymia kannattaa laskea ja intensiteettiä ei. Herkistelyn pituus on hyvä olla 8 - 14 päivän välissä ja harjoitustiheyttä ei kannata laskea. Sopiva voluumin lasku näyttäisi olevan noin puolet normaalista. Tämä riittää säilyttämään saavutetut ominaisuudet, mutta poistamaan väsymyksen.
TAULUKKO 5: Herkistelyn vaikuttaa
suorituskykyyn merkitsevästi meta-analyysin mukaan. N=koehenkilöiden määrä ja
P=tilastollinen merkitsevyys (tilastollisesti merkitsevimmät tulokset
lihavoitu). Tehokkain tapa herkistellä on laskea harjoitusvolyymia 41-60 % mutta
samalla pitää intensiteetti ylhäällä. Kestoltaan 8-14 vuorokauden
progressiivinen herkistely näyttäisi olevan pituudeltaan tehokkain. Mukailtu
(Bosquet et al., 2007).
Lähteet:
Auvinen, M. 2011. SUL koulutusohjeet 2011-2013. s. 58-70. Koonnut Rajala, T. 2011. Viitattu 3.11.2016.
Bompa, T. & Haff, G. 2009. Periodization - Theory and methodology of training. 5.
painos. Human Kinetics.
Bosquet, L.,
Montpetit, J., Arvisais, D. & Mujika I. 2007. Effects of tapering on
performance: A Meta-analysis. Medicine & Science in Sports
& Exercise, 39 (8), 1358-1365.
Häkkinen
& Ahtiainen. 2016. Maksimivoimaharjoittelu. Teoksessa Mero, A., Nummela,
A., Kalaja, S. & Häkkinen, K. (toim) Huippu-urheiluvalmennus. 1.painos. VK-Kustannus Oy, 250-265.
Krieger, J. 2010.
Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy: A
Meta-Analysis. Journal of strength and conditioning research, 24 (4),
1150-1159.
Mero, A., Nummela,
A., Keskinen, K. & Häkkinen, K. 2007. Urheiluvalmennus.
2.painos. VK-kustannus.
Rooney, K., Herbert, R. & Balnave, R. 1994. Fatigue contributes to the strength training stimulus.
Medicine
and science in sports and exercise, 26 (9), 1160-1164.
Räntilä, A. 2015. Yleisurheilun heittolajien fyysiset
suorituskykyvaatimukset käytännön kenttätesteissä ja niiden vaikutus
kilpailutuloksiin. Haaga-Helia, opinnäytetyö.
Schoenfeld, B.
2013. Potential mechanisms for a role of metabolic stress in hypertrophic
adaptations to resistance training. Sports medicine, 43 (3), 179-194.
Schiaffino, S., Dyar, K., Ciciliot, S., Blaauw, B. &
Sandri, M. 2013. Mechanisms regulating skeletal muscle growth and atrophy. Federation of European Biochemical
Societies, 280,
4294–4314.
Schott, J.,
McCully, K. & Rutherford, O. 1995. The role of metabolites in strength
training. II. Short versus long isometric contractions. European journal of
applied physiology and occupationally physiology, 71 (4), 337-341.
Yrjölä, M. 2000. Suomalainen Kuulantyöntö. Lahtiprint
- Lahti.
Zatsiorsky, V. & Kraemer,
W. 2006. Science and Practice of Strength Training. 2. painos. Human Kinetics.
Aapo Räntilä
Olen pääkaupunkiseudulla vaikuttava voima-, fysiikka- ja yleisurheiluvalmentaja. Valmentajana olen toiminut kohta 10 vuoden ajan. Olen erityisen kiinnostunut urheilijoiden suorituskyvystä ja harjoittelun yksilöllisistä eroista urheilijoilla sekä periodisaatiosta. Saan urheilijat suoriutumaan lajissaan paremmin.
Tämä sivusto on luotu aputyökaluksi valmentajille, jotka haluavat perustaa valmentamisensa tieteelliseen näyttöön. Alalla on vielä paljon käytössä vanhentuneita toimintatapoja, jotka heikentävät urheilijoiden suorituskykyä. Pyrin jakamaan tieteellistä tietoa ja auttamaan objektiivisesti kaikkia valmentajia.