Kuukautiskierron vaikutus urheilusuorituksiin on monisyinen, mutta tieteellinen näyttö suorituskyvyn vaihtelusta eri vaiheissa on ristiriitaista.
Harjoittelua ei kannata liikaa periodisoida pitkällä aikavälillä kuukautiskierron mukaan.
Hormonaalinen ehkäisy voi lievittää kierron oireita, mutta sen vaikutus suorituskykyyn on usein neutraali tai hieman negatiivinen.
Korkealaatuista tutkimusta tarvitaan lisää yksilöllisten erojen ja lyhytaikaisten hormonaalisten vaihteluiden vaikutusten ymmärtämiseksi.
Energiavaje ja kuukautiskierron häiriöt, kuten amenorrea, voivat merkittävästi haitata terveyttä ja suorituskykyä sekä selittää kuukautiskierron aikaista suorituskyvyn laskua.
Valmentajien ja urheilijoiden välinen avoin keskustelu sekä moniammatillinen tuki ovat avainasemassa kuukautiskierron vaikutusten hallinnassa.
Lukuaika: n. 7–8 minuuttia
Kuukautiskierron vaikutukset urheilusuorituksiin
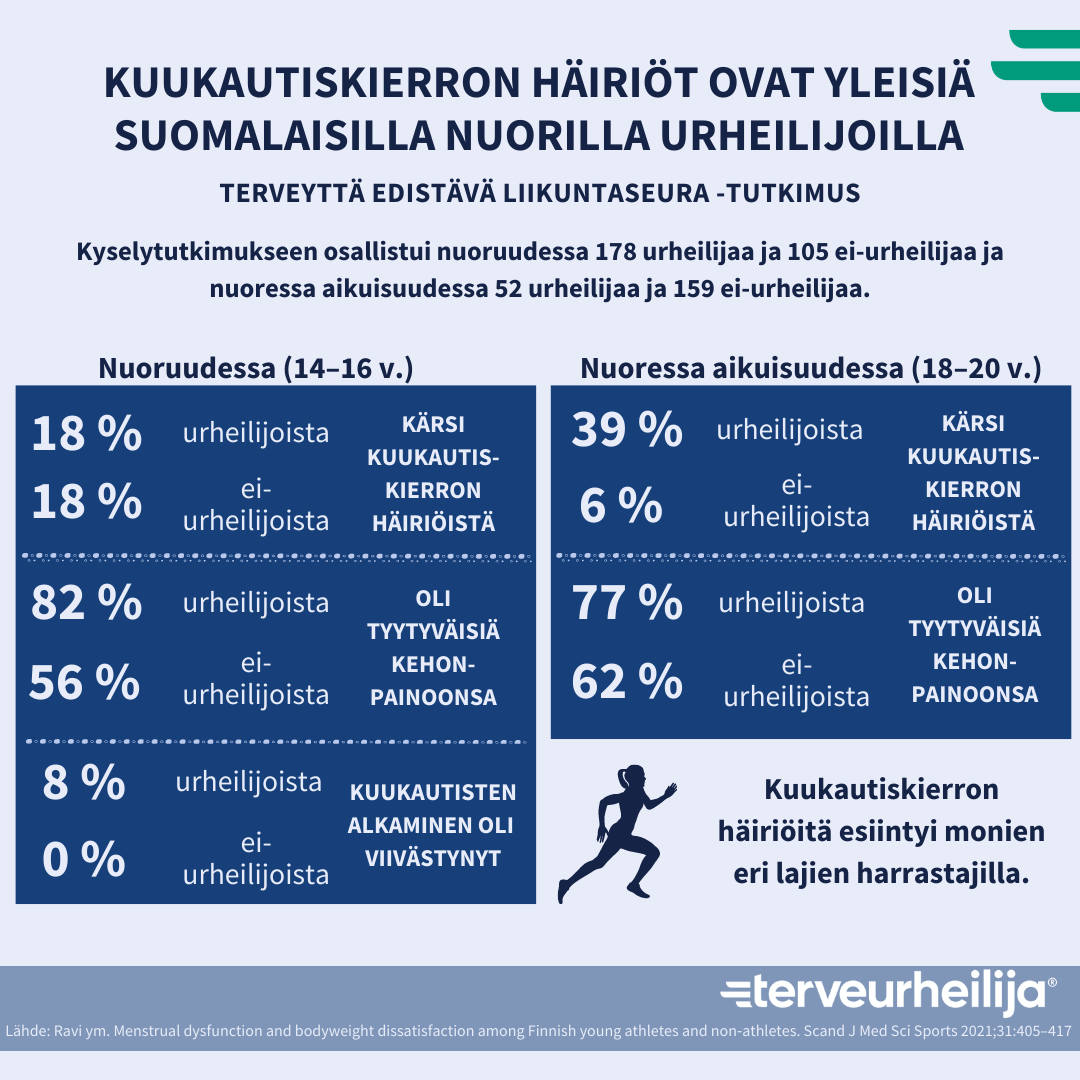
Urheilevilla naisilla havaitaan usein kuukautishäiriöitä, joita ovat muun muassa ovulaation puuttuminen, luteaalivaiheen lyheneminen sekä kuukautisten harventuminen tai niiden poisjääminen (amenorrea). On arvioitu, että yli puolet aktiivisesti liikkuvista ja urheilijanaisista kärsivät jonkinlaisesta kuukautiskiertoon liittyvästä häiriöstä. Näiden taustalla voi olla elimistön kuukautiskiertoa säätelevien hormonipitoisuuksien aleneminen, joka voi johtua esimerkiksi kulutukseen nähden alhaisesta energiansaannista, liiallisesta harjoittelusta tai kokonaiskuormituksesta ja/tai fyysisestä tai psyykkisestä stressistä. On ratkaisevan tärkeää, ettei kuukautiskierron häiriöitä pidetä normaalina, vaan niitä tutkitaan ja hallitaan moniammatillisesti koko urheilijan uran ajan. Usein esimerkiksi kuukautisten pois jäämistä pidetään normaalina osana kovaa harjoittelua, mutta tämä ei ole totta. Avoin keskustelu kuukautisista auttaa valmentautumisessa huomioimaan paremmin kuukautiskierron mahdolliset yksilölliset vaikutukset urheilijan suorituskykyyn.
Kuva 1: Kuukautiskierron häiriöt ovat hyvin yleisiä suomalaiilla nuorilla urheilijoilla. Lähde: Ravi, S., Waller, B., Valtonen, M., Villberg, J., Vasankari, T., Parkkari, J., Heinonen, O. J., Alanko, L., Savonen, K., Vanhala, M., Selänne H., Kokko, S. & Kujala, U.M. Menstrual dysfunction and body weight dissatisfaction among Finnish young athletes and non-athletes. Scandinavian Journal of Medicine and Science in Sports 2021;31:405–417. Infograafi: terveurheilija.fi
Naisurheilijan hormonitoiminta
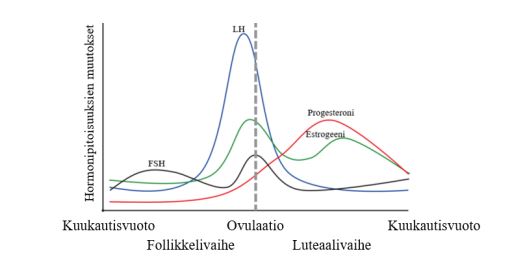
Kuukautiskierto jaetaan tavallisimmin kolmeen vaiheeseen hormonipitoisuuksissa tapahtuvien muutosten mukaan. Kaksi keskeistä kuukautiskiertoon liittyvää hormonia ovat estradioli (yksi estrogeenin esiintymismuodoista ja tässä tekstissä käytetään pitkälti estrogeeni-nimitiystä) ja progesteroni eli keltarauhashormoni. Estradiolin ja keltarauhashormonin pitoisuuksien vaihtelulla kuukautiskierron aikana on huomattavia vaikutuksia naisurheilijan fysiologiaan.
Androgeenihormoneihin lukeutuvan sukupuolihormonin testosteronin vaikutukset ovat erittäin hyvin tutkittuja, mutta estrogeenin ja progesteronin vaikutukset lihasmassan säätelyyn ovat heikommin tiedossa. Erilisissä tutkimuksissa on kuitenkin osoitettu estrogeenin vaikuttavan voimaharjoittelun adaptaatioihin, kuten proteiinin synteesiin ja myös degradaatioon, myosiinin toimintoihin sekäs satelliittisolujen aktiivisuuteen (18, 19). Näillä hormoneilla onkin suuri vaikutus voiman ja lihasmassan kehittymiseen pitkällä aikavälillä.
Lisäksi estrogeenin puute ja estrogeeni reseptorisignaloinnin vähentyminen vähentää satelliittisolujen määrää lihaksissa (19, 20), mikä saattaa vauhdittaa lihasmassan ja voiman vähentymistä esimerkiksi vaihdevuotisilla naisilla. Tämä tukee ajatusta siitä, että naisurheilijalla myös estrogeenillä on iso vaikutus voiman ja lihasmassan säätelyssä.
Kuukautiskierron suhteen estrogeenitasot ovat korkeimmillaan myöhäisessä follikulaarivaiheessa ennen ovulaatiota, kun taas progesteroni on korkeimmillaan ovulaation jälkeen puolessa välissä luteaalivaihetta. Tiedämme hormonien pienentymisen pitkällä aikavälillä olevan tuhoisaa lihas- ja voimaharjoittelun näkökulmasta, mutta kuukautiskierto vaikuttaa hormoneihin vain muutamien päivien ajan. Onko tällä lyhytaikaisella hormonivaihtelulla merkittävää vaikutusta akuuttiin suorituskykyyn tai pitkäaikaisiin hermolihasjärjestelmän adaptaatioihin?
Kuva 2: Kuukautiskierron vaiheet ja hormonipitoisuuksien muutokset. Ensimmäinen kierron vaiheista on follikkelivaihe, toinen vaihe on ovulaatio, eli munasolun irtoaminen ja kolmas vaihe luteaalivaihe. Estradiolipitoisuus saavuttaa yleensä huippupitoisuutensa juuri ennen ovulaation alkua.
Subjektiivisesti kuukautiskierto vaikuttaa suorituskykyyn, muttei objektiivisesti?
Vaikka urheilijat itse kokevat kuukautiskierron vaikuttavan suorituksiinsa, objektiiviset tutkimustulokset eivät aina tue tätä käsitystä. Esimerkiksi 241 urheilijan tutkimuksessa ei havaittu suorituskyvyn eroja eri kierron vaiheissa (6).
Hormonitoiminnalla on selitetty suorituskyvyn vaihtelua
Estrogeenihormonia on pidetty keskeisenä tekijänä follikulaarisen vaiheen suorituskyvyn nousussa, kun taas keltarauhashormoni eli progestiini yhdistetään luteaalivaiheen heikentyneeseen suorituskykyyn (7). Progestiini nostaa kehon lämpötilaa, mikä voi haitata aineenvaihduntaa ja aiheuttaa lisärasitusta sydän- ja verisuonijärjestelmälle (8). Estrogeeni puolestaan tehostaa glukoosin ottoa lihakseen supistuksen jälkeen, kun taas progestiini saattaa estää tätä prosessia (9). Lisäksi estrogeeni edistää vapaiden rasvahappojen käyttöä energialähteenä, mikä säästää lihasten glykogeenivarastoja liikunnan aikana (7).
Lyhyesti yksi estrogeenin ilmentymismuodon, estradiolin, vaikutus suorituskykyyn on merkittävä. Estradioli voi edistää lihaskudoksen rakentumista, voimantuottoa ja aineenvaihduntaa säästämällä lihasten energiavarastoja sekä lisäämällä hiilihydraattien käyttöä. Sillä on myös antioksidanttisia ja tulehdusta vähentäviä vaikutuksia, jotka suojaavat lihaskudosta vaurioilta. Lisäksi estradioli voi parantaa hermoston toimintaa ja lihasten tahdonalaista aktivointia.
Olisi loogista olettaa, että suorituskyky vaihtelee kuukautiskierron vaiheiden mukaan, erityisesti silloin, kun estrogeenitasot ovat alhaalla – esimerkiksi myöhäisessä luteaalivaiheessa tai aikaisessa follikulaarivaiheessa. Kuitenkin suorituskyvyn on havaittu olevan yllättävän vakaa kierron eri vaiheissa.
Harjoittelun ohjelmointi kuukautiskierron mukaan
Harjoittelun ohjelmointia kuukautiskierron mukaan tukee vain muutama tutkimus. Näistä tarkemmin vain kaksi artikkelia on raportoinut, että harjoittelun keskittäminen follikulaariseen vaiheeseen, jolloin harjoitusfrekvenssi ja -volyymi ovat korkeampia, tuottaa parempia tuloksia verrattuna luteaalivaiheen painottamiseen tai tasaisesti kierron eri vaiheisiin jaettuun harjoitteluun (24, 25). Laajemmin tarkasteltuna tulokset eivät kuitenkaan ole yksiselitteisiä. Uudessa katsauksessa (26), jossa huomioitiin kaikki saatavilla oleva data, tulokset vaihtelivat merkittävästi. Suuri osa tutkimuksista oli metodologisesti heikkotasoisia, eikä niiden perusteella voida tehdä luotettavia johtopäätöksiä harjoittelun ohjelmoinnin hyödyistä kuukautiskierron mukaan. Nykyisen tiedon perusteella kuukautiskierron mukaisella harjoittelulla ei ole pitkän aikavälin vaikutusta voiman, suorituskyvyn tai lihaskasvun kehitykseen.
Mutta miksi tässä on niin räikeä ero subjektiivisesti koetun ja objektiivisesti mitattujen tekijöiden välillä?
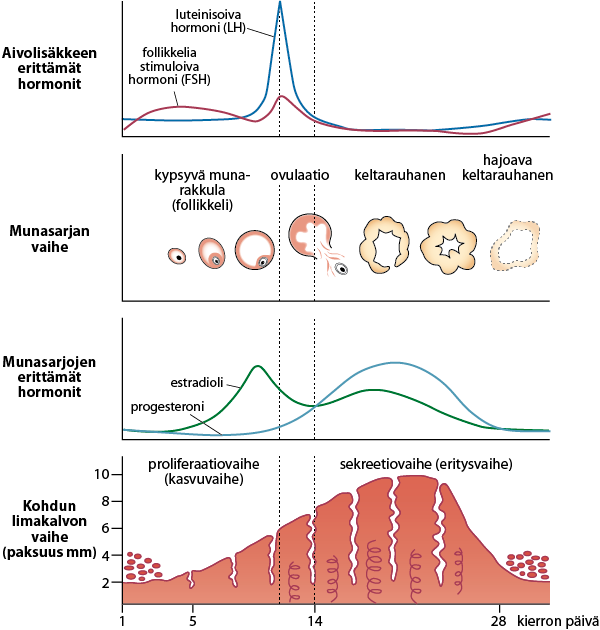
Kuva 3: Hypotalamus-aivolisäke-munasarja-akselin normaali toiminta. Lähde: Luiro-Helve ja Holopainen. Lääketieteellinen Aikakauskirja Duodecim 2023;139(5):359-367
Metodologia ei huomioi yksilöllisiä eroja
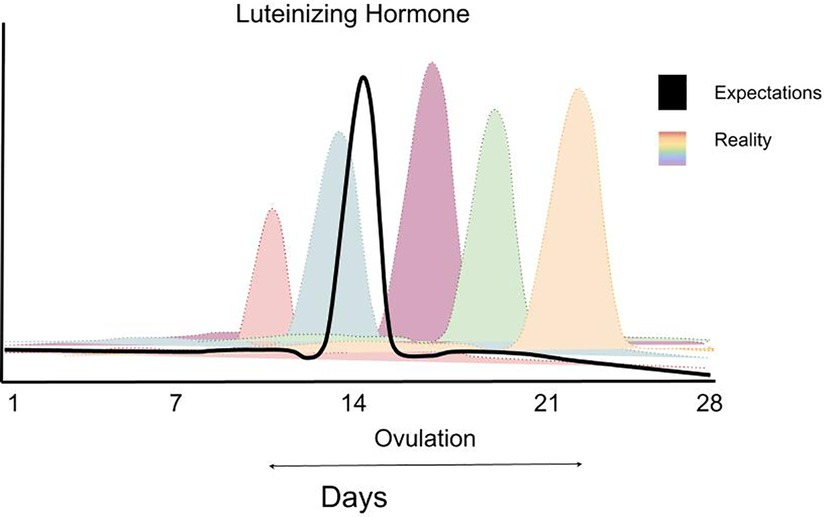
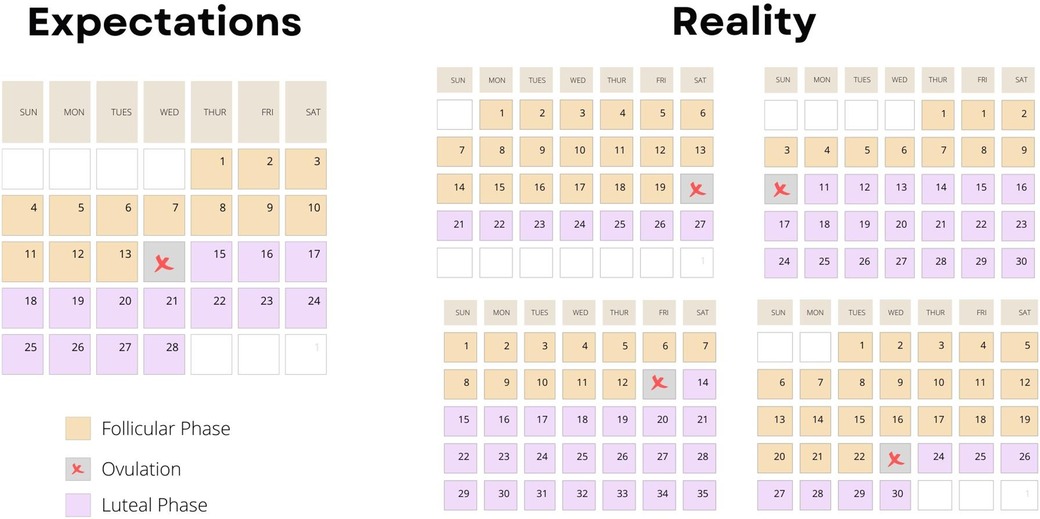
Kuukautiskierron vaiheen määrittämisen haasteet korostuvat tutkimusten ristiriitaisissa tuloksissa. Follikulaarisen vaiheen kesto voi vaihdella 10–22 päivän välillä ja luteaalivaiheen 7–17 päivän välillä (27) (katso kuva 4). Jo vuonna 1926 Ludvig Fränkel totesi, että kuukautiskierron ainoa säännöllisyys on epäsäännöllisyys (28). Tästä huolimatta naisten kuukautiskierron oletetaan yhä olevan tasainen 28 päivän sykli, jossa ovulaatio tapahtuu kolmantenatoista päivänä (kuva 5). Esimerkiksi luteinisoivan hormonin monitorointi, yksi yleisimmin käytetyistä kierron vaiheiden määrittämismenetelmistä, voi osoittaa ovulaation yhden päivän tai 2–6 päivän ajalta riippuen yksilöstä (29).
Kuva 4: Kaaviokuva, joka havainnollistaa luteinisoivan hormonin piikin vaihtelua naisilla ja korostaa sitä, kuinka epätarkoituksenmukaista on suunnitella suurempaa harjoitusvolyymia kuukautiskierron tietylle vaiheelle. Tiedot ovat teoreettisia, mutta perustuvat havaintoihin, jotka osoittavat LH-piikin huomattavaa vaihtelua naisilla. Lähde: 26.
Sarwar et al. (1996) (10) havaitsivat, että voima kasvaa ovulaation aikana, mikä yhdistettiin estrogeenitasojen nousuun. Näitä tuloksia ei kuitenkaan ole pystytty toistamaan muissa tutkimuksissa, kuten penkkipunnerruksen maksimitoistoissa tai aerobisessa suorituskyvyssä (11). Tuoreessa meta-analyysissa (12) havaittiin vähäinen suorituskyvyn heikkeneminen varhaisessa follikulaarivaiheessa, mutta tutkimusten vähäinen määrä ja laatu tekevät johtopäätöksistä epävarmoja. Toinen systemaattinen katsaus (13), jossa tutkittiin 314 eliittiurheilijaa, ei löytänyt todisteita kuukautiskierron vaikutuksesta suorituskykyyn. Toisaalta vastakkaisiakin tuloksia on, jossa huomattiin että varhainen follikulaarivaihe on epäedullinen kaikille voimatyypeille. Isometrinen voima on huipussaan myöhäisessä follikulaarivaiheessa, kun taas isokineettinen voima saavuttaa huippunsa ovulaation aikana. Dynaaminen voima on optimaalisimmillaan myöhäisessä follikulaarivaiheessa (30).
On selvää, että itsearvioidun kuukautiskierron vaikutusten ja todellisten fysiologisten suorituskykyerojen välillä on eroja. Tarvitaan lisää korkealaatuista tutkimusdataa, jotta voidaan ymmärtää kuukautiskierron vaiheen ja suorituskyvyn monimutkaista vuorovaikutusta.
Kuva 5: Havainnollistava kuva, joka osoittaa kuinka epätarkoituksenmukaista on suunnitella suurempaa harjoitusvolyymia kuukautiskierron tietylle vaiheelle, jos oletetaan ovulaation ajankohdan tapahtuvan aina samassa vaiheessa kuukautta. Lähde: 26.
Mutta onko kyseessä kuukautiskierron aiheuttama suorituskyvyn lasku vai laiskaa valmennusta?
Kuukautiskierron häiriöt voivat johtua useista tekijöistä, joista yksi merkittävimmistä on krooninen energiavaje. Tämä voi johtaa hypotalamisesta amenorreaan, jossa kuukautiset jäävät väliin hypotalamuksen toimintahäiriön seurauksena (3). Hypotalaminen amenorrea on huomattavan yleinen erityisesti kestävyysjuoksijoilla, joista jopa 65 % kärsii tästä tilasta, sekä balettitanssijoilla, joiden kohdalla esiintyvyys on jopa 79 %. Yleisväestössä vastaava esiintyvyys on vain noin 5 % (4, 5). Tämä viittaa siihen, että kuukautiskiertoon liittyvät suorituskyvyn haasteet eivät välttämättä johdu pelkästään kierron fysiologisista vaiheista, vaan taustalla voi olla muita tekijöitä, kuten energiatasapainon häiriöt, jotka ovat hallittavissa asianmukaisilla toimenpiteillä ja tuella. Toinen merkittävä tekijä on runsaat kuukautisvuodot, jotka voivat altistaa raudan puutokselle. Raudalla on tärkeä rooli elimistön hapenkuljetuksessa ja sen puute heikentää suorituskykyä.
Onko ehkäisytavalla väliä suorituskyvyn näkökulmasta?
Pyrkimyksenä lieventää kehon sisäisten sukupuolihormonien vaihtelun mahdollisia fysiologisia vaikutuksia – tai ainakin vähentää heikomman suorituskyvyn kokemusta kuukautiskierron aikana – urheilijat saattavat käyttää hormonaalista ehkäisyä kierron hallitsemiseksi. Hormonaalista ehkäisyä hyödynnetään usein kuukautisten alkamisen viivästyttämiseen tietyn kilpailupäivän yli, kierron oireiden lievittämiseen tai esimerkiksi runsaan kuukautisvuodon aiheuttaman raudanpuuteanemian ehkäisyyn. Joissakin tapauksissa urheilijat voivat trisyklisellä lähestymistavalla välttää kuukautisten tyhjennysvuodon koko kilpailukauden ajan.
Hormonaalisen ehkäisyn vaikutukset suorituskykyyn
Vaikka hormonaalista ehkäisyä käytetään urheilijoiden keskuudessa laajasti, tutkimukset eivät ole osoittaneet sen parantavan suorituskykyä merkittävästi. Jonesin ym. (31) kyselytutkimuksen mukaan suorituskyvyn koettu parantuminen oli vähäistä hormonaalista ehkäisyä käyttävillä urheilijoilla. Lisäksi tuoreessa meta-analyysissa (14) todettiin, että hormonaalisen ehkäisyn käyttö voi aiheuttaa pienen mutta todennäköisesti negatiivisen vaikutuksen urheilusuoritukseen. Esimerkiksi ehkäisypillereiden käytöllä ei ole havaittu positiivisia vaikutuksia lihasvoimaan verrattuna ei-hormonaaliseen ehkäisyyn (15), ja se voi jopa laskea maksimaalista hapenottokykyä (16).
Näiden tulosten perusteella hormonaalisen ehkäisyn käyttöä ei voida suositella suorituskyvyn optimoimisen näkökulmasta. Toisaalta ehkäisyn käytöllä voi olla tärkeitä sosiaalisia ja henkisiä hyötyjä, jotka ovat merkityksellisiä suorituskyvyn ulkopuolella. Esimerkiksi Jonesin (31) tutkimuksessa 81 % ei-hormonaalista ehkäisyä käyttävistä raportoi kipeitä kuukautiskipuja, kun taas hormonaalista ehkäisyä käyttävistä vain 53 % raportoi samanlaisia oireita. Lisäksi jotkut tutkimukset ovat osoittaneet, että hormonaaliset ehkäisymenetelmät eivät välttämättä vaikuta suorituskykyyn merkittävästi (17).
Minipillerit ja urheilijat
Pelkkää progestiinia, eli keltarauhashormonia, sisältävät ehkäisyvalmisteet, kuten minipillerit ja pitkävaikutteiset menetelmät (esim. kapselit ja kierukat), ovat kasvattaneet suosiotaan viime vuosina. Näillä valmisteilla on monia etuja, kuten se, että ne eivät lisää veritulppariskiä. Mutta progestiinipohjaisten menetelmien mahdollinen negatiivinen vaikutus suorituskykyyn pitkällä aikavälillä on selkeästi osoitettu ja huippu-urheilussa pienikin myönnytys kehittymisen suhteen voi olla ratkaiseva. Keltarauhashormonilla on esimerkiksi monilta osin estradiolin toimintaa hillitsevä vaikutus. Tällä hetkellä ei kuitenkaan ole riittävästi tutkimustietoa suositella mitään tiettyä ehkäisymenetelmää yleisesti huippu-urheilijoille. Siksi urheilijoiden kannattaa keskustella aiheesta paitsi terveydenhuollon ammattilaisten myös valmentajiensa kanssa löytääkseen itselleen parhaiten sopivan ratkaisun.
Yhteenveto
Kuukautiskierron mukainen harjoittelun ohjelmointi ei toimi, eikä sitä kannata käyttää. Tulosten puute johtuu luultavasti metodologisista ongelmista ja kuukautiskiertojen eri vaiheiden määrittelyn vaikeudesta. Ennen kuin lisää kattavaa dataa saadaan, niin en hukkaisi aikaa tähän. Kuukautiskierron mukainen harjoittelun ohjelmointi ei ole näyttöön perustuva lähestymistapa nykyisellä tietämyksellämme. Laadukkaan näytön puuttuessa kuukautiskiertoon perustuvien harjoitusohjelmien räätälöimisessä valmentajien ja urheilijoiden tulisi rakentaa ohjelmointi yksilön mukaan. Kuukautiskierron vaikutus voi olla huomioitava tekijä ohjelmaa suunniteltaessa, samoin kuin monet muut tutut tekijät, kuten: ravitsemus, väsymys, unen laatu, stressi, loukkaantumiset, motivaatio ja muut harjoittelun sisäiset muuttujat. Myös kuukautisoireet voivat vaikuttaa joidenkin naisten harjoitussuoritukseen merkitsevästi. Tämän takia urheilijan omaa tuntemusta kannattaa kuunnella herkällä korvalla.
Lähteet:
1. Costello JT, Bieuzen F, Bleakley CM. Where are all the female participants in sports and exercise medicine research? Eur J Sport Sci. (2014) 14(8):847–51. doi: 10.1080/17461391.2014.911354
2. Cowley ES, Olenick AA, McNulty KL, Ross EZ. “Invisible sportswomen”: the sex data gap in sport and exercise science research. Women Sport Physical Activity J. (2021) 29(2):146–51. doi: 10.1123/wspaj.2021-0028
3. Jones BP, L’Heveder A, Saso S, Yazbek J, Smith JR, Dooley M. Sports gynaecology. Obstet Gynaecol. (2019) 21(2):85–94. doi: 10.1111/tog.12557
4. Dusek T. Influence of high intensity training on menstrual cycle disorders in 56 athletes. Croat Med J. (2001) 42:79–82.11172662
5. Abraham SF, Beumont PJV, Fraser IS, Llewellyn-Jones D. Body weight, exercise and menstrual status among ballet dancers in training. Br J Obstet Gynaecol. (1982) 89:507–10. doi: 10.1111/j.1471-0528.1982.tb03649.x
6. Kishali NF, Imamoglu O, Katkat D, Atan T, Akyol P. Effects of menstrual cycle on sports performance. Int J Neurosci. (2006) 116(12):1549–63. doi: 10.1080/00207450600675217
7. Lebrun CM, Joyce S, Constantini NW. Effects of female reproductive hormones on sports performance. In: Constantini N, Hackney A, editors. Endocrinology of Physical Activity and Sport. Contemporary Endocrinology. Totowa: Humana Press (2013). p. 267–302.
8. Janse de Jonge XA. Effects of the menstrual cycle on exercise performance. Sports Med. (2003) 33(11):833–51. doi: 10.2165/00007256-200333110-00004
9. Oosthuyse T, Bosch AN. The effect of the menstrual cycle on exercise metabolism: implications for exercise performance in eumenorrhoeic women. Sports Med. (2010) 40(3):207–27. doi: 10.2165/11317090-000000000-00000
10. Sarwar R, Niclos BB, Rutherford OM. Changes in muscle strength, relaxation rate and fatiguability during the human menstrual cycle. J Physiol. (1996) 493(Pt 1):267–72. doi: 10.1113/jphysiol.1996.sp021381
11. Quadagno D, Faquin L, Lim G-N, Kuminka W, Moffatt R. The menstrual cycle: does it affect athletic performance? Phys Sportsmed. (1991) 19(3):121–4. doi: 10.1080/00913847.1991.11702172
12. McNulty KL, Elliott-Sale KJ, Dolan E, Swinton PA, Ansdell P, Goodall S, et al. The effects of menstrual cycle phase on exercise performance in eumenorrheic women: a systematic review and meta-analysis. Sports Med. (2020) 50(10):1813–27. doi: 10.1007/s40279-020-01319-3
13. Meignié A, Duclos M, Carling C, Orhant E, Provost P, Toussaint JF, et al. The effects of menstrual cycle phase on elite athlete performance: a critical and systematic review. Front Physiol. (2021) 12:654585. doi: 10.3389/fphys.2021.654585
14. Elliott-Sale KJ, McNulty KL, Ansdell P, Goodall S, Hicks KM, Thomas K, et al. The effects of oral contraceptives on exercise performance in women: a systematic review and meta-analysis. Sports Med. (2020) 50(10):1785–812. doi: 10.1007/s40279-020-01317-5
15. Nichols AW, Hetzler RK, Villanueva RJ, Stickley CD, Kimura IF. Effects of combination oral contraceptives on strength development in women athletes. J Strength Cond Res. (2008) 22(5):1625–32. doi: 10.1519/JSC.0b013e31817ae1f3
17. Elliott-Sale, K. J., Mcnulty, K. L., Ansdell, P., Goodall, S., Hicks, K. M., Thomas, K., Swinton, P. A., & Dolan, E. (2020). The effects of oral contraceptives on exercise performance in women: A systematic review and meta-analysis. Sports Medicine (Auckland, N.Z.), 50(10), 1785–1812.
18. Chidi-Ogbolu N BK. Effect of estrogen on musculoskeletal performance and injury risk. Front Physiol. (2019) 9:1834. 10.3389/fphys.2018.01834
19. Collins BC, Arpke RW, Larson AA, Baumann CW, Xie N, Cabelka CA, et al. Estrogen regulates the satellite cell compartment in females. Cell Rep. (2019) 28:368–81.
20. Ikeda K, Horie-Inoue K, Inoue S. Functions of estrogen and estrogen receptor signaling on skeletal muscle. J Steroid Biochem Mol Biol. (2019) 191:105375. 10.1016/j.jsbmb.2019.105375
21. Chidi-Ogbolu N BK. Effect of estrogen on musculoskeletal performance and injury risk. Front Physiol. (2019) 9:1834. 10.3389/fphys.2018.01834
22. Collins BC, Arpke RW, Larson AA, Baumann CW, Xie N, Cabelka CA, et al. Estrogen regulates the satellite cell compartment in females. Cell Rep. (2019) 28:368–81.
23. Ikeda K, Horie-Inoue K, Inoue S. Functions of estrogen and estrogen receptor signaling on skeletal muscle. J Steroid Biochem Mol Biol. (2019) 191:105375. 10.1016/j.jsbmb.2019.105375
24. Sung E, Han A, Hinrichs T, Vorgerd M, Manchado C, Platen P. Effects of follicular versus luteal phase-based strength training in young women. SpringerPlus. (2014) 3:668. 10.1186/2193-1801-3-668
25. Wikström-Frisén L, Boraxbekk CJ, Henriksson-Larsén K. Effects on power, strength and lean body mass of menstrual/oral contraceptive cycle-based resistance training. J Sports Med Phys Fitness. (2017) 57:43–52. 10.23736/S0022-4707.16.05848-5
26. Colenso-Semple LM, D’Souza AC, Elliott-Sale KJ, Phillips SM. Current evidence shows no influence of women’s menstrual cycle phase on acute strength performance or adaptations to resistance exercise training. Front Sports Act Living. 2023 Mar 23;5:1054542. doi: 10.3389/fspor.2023.1054542. PMID: 37033884; PMCID: PMC10076834.
27. Shea A, Vitzthum V. The extent and causes of natural variation in menstrual cycles: integrating empirically-based models of ovarian cycling into research on women’s Health. Drug Discov Today Dis Models. (2020) 32:41–9. 10.1016/j.ddmod.2020.11.002¨
28. Vitzthum V. Field methods and strategies for assessing female reproductive functioning. Am J Hum Biol. (2021) 33:e23513. 10.1002/ajhb.23513
29. Su HW, Yi YC, Wei TY, Chang TC, Cheng CM. Detection of ovulation, a review of currently available methods. Bioeng Transl Med. (2017) 2(3):238–46. 10.1002/btm2.10058
30. Niering M, Wolf-Belala N, Seifert J, Tovar O, Coldewey J, Kuranda J, Muehlbauer T. The Influence of Menstrual Cycle Phases on Maximal Strength Performance in Healthy Female Adults: A Systematic Review with Meta-Analysis. Sports. 2024; 12(1):31. https://doi.org/10.3390/sports12010031
31. Jones, B. P., L’Heveder, A., Bishop, C., Kasaven, L., Saso, S., Davies, S., Chakraverty, R., Brown, J., & Pollock, N. (2024). Menstrual cycles and the impact upon performance in elite British track and field athletes: a longitudinal study. Frontiers in sports and active living, 6, 1296189. https://doi.org/10.3389/fspor.2024.1296189
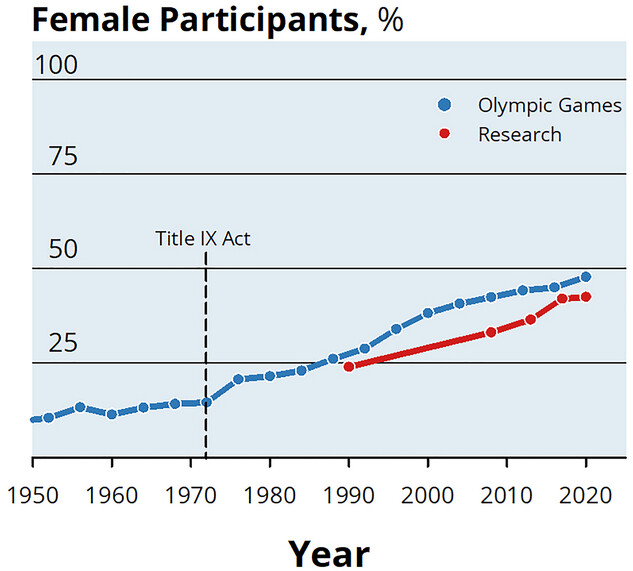
Pariisin Olympialaisissa oli ensimmäistä kertaa miesten ja naisten suhde tasan 50 % urheilijoiden kokonaismäärästä. Aikanaan vielä Rooman olympialaisissa vuonna 1960 urheilijoista vain 11 % oli naisia. Tämä nouseva trendi urheilussa ei ole vielä ihan samalla tavalla näkynyt tutkimuksessa ja meillä on vielä isojakin aukkoja ymmärryksessämme naisten fysiologiassa. Arvioiden mukaan noin kolmannes liikuntatutkimukseen osallistujista on naisia (1). Ja esimerkiksi vuosina 2014-2020 vain 6% liikuntatutkimuksista oli tehty naisille, kun sama luku miehille oli 31% (2). Tutkimus tulee yleisesti ajallisesti perässä käytäntöä tutkimuksen toteuttamiseen liittyvien ajankäytöllisten syitten takia, joten tulemme tulevaisuudessa näkemään enemmän naisiin liittyviä tutkimuksia. Esimerkiksi naisten suhteellinen osuus tutkimuksissa kasvoi vuoden 1991 22 prosentista jopa 36 prosenttiin vuoteen 2021 mennessä (3).
Miehiä voidaan yleisesti kuvata olympialaisten perinteisen latinalaisen moton, citius – altius – fortius, mukaisesti, joka kääntyy ‘nopeammin, korkeammalle, voimakkaammin’. Miehet, erityisesti huippu-urheilijat, ovat nopeampia sekä lyhyillä että pitkillä matkoilla kaikissa liikkumisen muodoissa, tuottavat enemmän lihasvoimaa ja pystyvät siten hyppäämään korkeammalle, ja ovat fyysisesti vahvempia kuin naiset (2). Miten naisurheilijaa pitäisi sitten harjoittaa? Eroaako harjoittelu miesten harjoittelusta?
KUVA 1: Naisten osuus urheilun parissa on kasvanut tasaisesti ja lähes samaa vauhtia on kasvanut naisiin liittyvä tutkimus. Kuvasta puuttuu Pariisin Olympialaiset vuonna 2024, jossa saavutettiin 50% suhde urheilevien miesten ja naisten välillä. Kuva: Hunter et al. (1).
Kuinka isot sukupuolten väliset erot ovat?
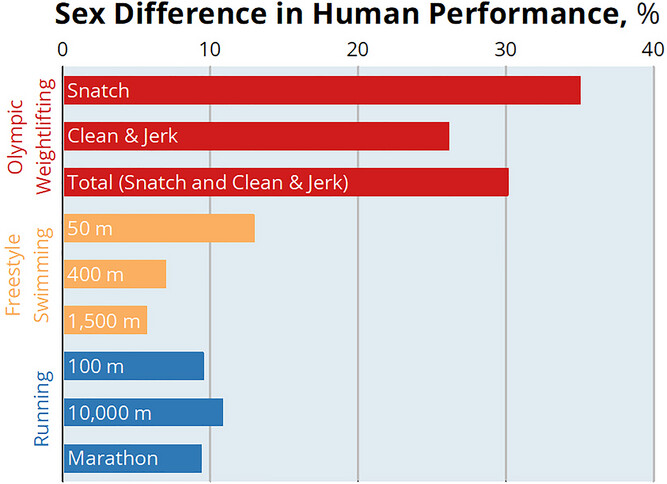
Maailmanennätykset urheilutapahtumissa ovat hyviä merkkejä kertomaan sukupuolieroista suorituskyvyssä. Ennätykset heijastelevat sitä, missä on suurimmat sukupuolten väliset erot ihmisten suorituskyvyssä. Koska lähtökohtaisesti kaikki maailmanennätyksen tehneet urheilijat ovat harjoitelleet erittäin pitkään ja laadukkaasti, joten erot muodostuvat fysiologiasta ja anatomiasta.
Miehet ylittävät naiset noin 5–35 prosentilla, vaihdellen urheilutapahtuman fysiologisten vaatimusten mukaan. Yleisesti ottaen sukupuolten väliset erot huippu-urheilusuorituksessa ovat suurimmat lajeissa, jotka liittyvät tiiviimmin maksimaaliseen lihasvoimaan tai -tehoon, kuten painonnostossa. Sen sijaan sukupuolten väliset erot suorituskyvyssä ovat pienempiä, vaikka edelleen merkittäviä, tapahtumissa, jotka liittyvät enemmän aerobiseen tehoon ja lihasten oksidatiiviseen kapasiteettiin, kuten maratonjuoksussa ja pitkänmatkan uinnissa.
KUVA 2: Isoimmat erot naisten ja miesten välillä löytyvät räjähtävää voimaa vaativista urheilulajeista, kuten painonnostosta ja pienimmät aerobisia ominaisuuksia vaativista lajeista. Kuva: Hunter et al. (1).
Mikä erottaa naiset miehistä fysiologisesti?
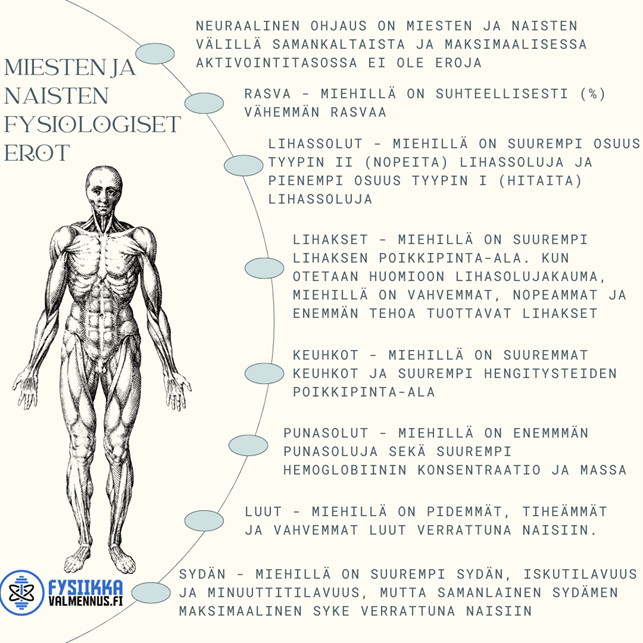
Erot muodostuvat lihaksesta, ei hermostosta
Sukupuolten välisten erojen mekanismit maksimaalisessa voimassa ja tehossa ovat pääasiassa lihaskudoksesta peräisin, ja hermoston ohjauksessa on vain vähäisiä eroja. Erilaiseen luustolihaksen morfologiaan ja koostumukseen vaikuttaa pääasiassa geenien ilmentyminen ja sukupuolispesifit hormonit, kuten testosteroni. (5,6.)
Yksittäisten lihassolujen absoluuttinen määrä ei kuitenkaan eroa sukupuolten välillä (7). Lisäksi yksittäisen lihassolun jännitys on samanlainen sukupuolten välillä niin nuorilla kuin vanhemmilla aikuisilla (8, 9). Näin ollen miesten suurempi lihasvoima verrattuna naisiin selittyy suuremmalla lihasmäärällä, ei eroista yksittäisessä lihassolussa.
KVUA 3: Miesten ja naisten fysiologiset erot tiivistettynä kuvaan. Kuvan lähde Instagramista.
Testosteronin merkitys
Testosteroni on miesten ensisijainen sukupuolihormoni ja se on anabolinen steroidi. Sitä pidetään yhtenä tärkeimpänä selittäjästä sukupuolten välisissä eroissa ihmisen motorisessa suorituskyvyssä murrosiästä aikuisuuteen. Murrosiän ohittaneilla miehillä on jopa 15-kertaiset testosteronitasot naisiin nähden, ikään katsomatta. Tämä sekä kroonisen altistuksen fysiologiset ja anatomiset vaikutukset johtavat miehillä urheilulliseen etuun verrattuna naisiin. (2.)
Testosteroni on siis huomattavan tärkeä tekijä miesten ja naisten välisissä eroissa, muttei kuitenkaan ainoa tekijä. Nuorten, terveiden miesten ryhmässä, joilla on tasaisesti korkeat endogeeniset testosteronipitoisuudet, on näyttöä siitä, että lihasten androgeenireseptorien määrä on läheisemmässä yhteydessä luustolihasten hypertrofiaan voimaharjoittelun jälkeen kuin hormonipitoisuudet (4).
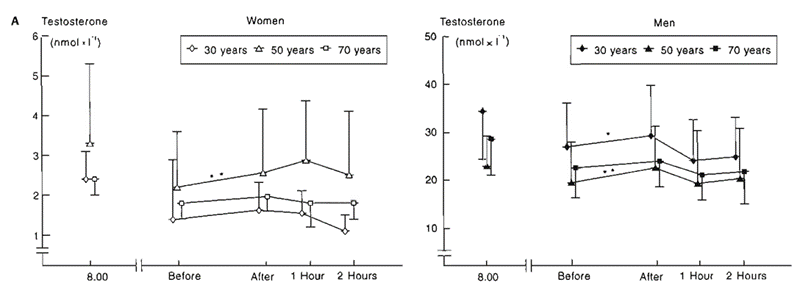
KUVA 4: Vasemmalla naisten testosteronivaste harjoitteluun ja oikealla miesten. After tarkoittaa reenin jälkeistä mittauspistettä. Kannattaa huomioida, että naisten mitta-asteikko on 0-6, kun miehillä se on 10-50. Mielenkiintoisena lisänä myös miten radikaalisti naisilla näyttää ikä vaikuttavan vasteisiin verrattuna miehiin, joilla vaste näyttää laskevan, muttei yhtä radikaalisti kuin naisilla. Lähde: Häkkinen, K., & Pakarinen, A. (1995). Acute hormonal responses to heavy resistance exercise in men and women at different ages. International journal of sports medicine, 16(8), 507–513. https://doi.org/10.1055/s-2007-973045
Miesten lihakset supistuvat nopeammin kuin naisten
Yksi tärkeä tekijä on myös lihasvoiman supistumisnopeus, mikä on miehillä nopeampaa kuin naisilla. Tämä johtuu pääasiassa siitä, että naisten kokonaislihaksessa on suhteellisesti suurempi osa tyypin I (hidas) lihassoluja ja pienempi osa tyypin II (nopeat) lihassoluja verrattuna miehiin (10). Lisäksi tyypin II (nopeat) lihassolut miehillä ovat suhteellisesti suurempia kuin naisilla.
Miesten suuremmassa kokonaislihaksessa on myös suhteellisesti enemmän tyypin II (nopeat) lihassolujen aluetta kuin naisilla. Myöskin myosiini-raskaanketjun (MHC) analyysi luustolihassoluista osoittaa, että naisilla on suurempi suhteellinen MHC I -pitoisuus kuin miehillä ja pienempi MHCII -pitoisuus (11). Mikä tarkoittaa suomeksi sitä, että tarkemmillakin analyyseillä on huomattu naisten omaavan enemmän hitaita lihassoluja kuin miesten. Mutta se ei kuitenkaan tarkoita, etteikö naiset voisi kasvattaa lihaksia.
Naisilla on sama potentiaali kasvattaa lihaksia kuin miehillä
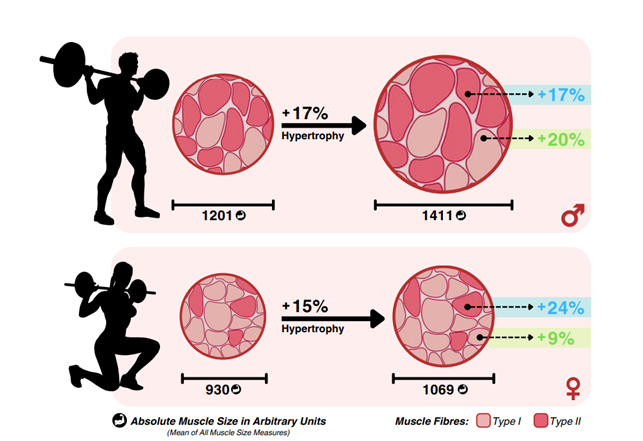
Vaikka naisilla on vähemmän lihasmassaa, niin naiset pystyvät kasvattamaan lihaksia ihan yhtä paljon kuin miehet. Absoluuttisesti miehillä kasvavat lihakset tietysti enemmän, mutta mielenkiintoisesti lihaksen kasvu on kuitenkin suhteellisesti samanlaista sukupuolten välillä. Eli rautalangasta vääntäen prosentuaalinen kasvu on samanlaista sukupuolten välillä. Lisäksi miehillä näyttäisi kasvavan tyypin I – lihassolut enemmän kuin naisilla, mutta tyypin II – lihassolujen kasvu on sukupuolten välillä samankaltaista toisin kuin usein ajatellaasn. (39.)
Sukupuolten välinen ero absoluuttisessa lihaskasvussa johtuu todennäköisesti lihasten lähtökoosta, kun taas suhteellinen lihaskasvu perustuu lähtökoon mukaiseen suhteelliseen kasvuun. Esimerkiksi, koska naisilla on keskimäärin vähemmän lihasmassaa lähtötilanteessa, absoluuttinen kasvu on pienempi, vaikka suhteellinen muutos olisi samanlainen kuin miehillä.
Sukupuolten välinen samankaltainen lihaskasvu tarkoittaisi, että sukupuolihormonien roolia lihaskasvussa aikuisiällä on saatetty yliarvioida. Muut fysiologiset tekijät voivat olla tärkeämpia, kuten mekaaniseen jännitykseen liittyvä anabolinen signalointi ja metabolinen stressi. Lisäksi voidaan pohtia estradiolin anabolisia ominaisuuksia, jotka voivat edistää lihaskasvua. Lisäksi äärimmäisen tärkeä positiivinen yhteys on lihaskasvun ja androgeenireseptorien määrän välillä. Näiden tekijöiden vaikutus harjoituksen jälkeisessä proteiinisynteesissä ja molekulaarisessa signaloinnissa, jotka käynnistävät lihaskasvun, voi olla tärkeämpi kuin itse hormoniprofiili. Toisaalta harjoituksen jälkeisellä anabolisten hormonien nousulla on vähintään pieni rooli lihasproteiinisynteesin stimuloimisessa. (39.)
KUVA 5: Lihaskasvu on suhteellisesti samanlaista sukupuolesta riippumatta. Miehillä kasvaa hiukan enemmän tyypin I – lihassolut ja tyypin II lihasolujen muutos on samankaltaista. Muokattu Refalo et al., (preprint) (39).
Naiset väsyvät vähemmän kuin miehet voimaharjoittelussa
Hermoston ohjauksen välillä ei ole eroja miesten ja naisten välillä. Samanlaiset ja lähes maksimaaliset tahdonalaisen aktivoinnin tasot miehillä ja naisilla osoittavat, että lihasmekanismit, eivät hermostolliset mekanismit, ovat ensisijaisesti vastuussa miesten suuremmasta voimantuotosta verrattuna naisiin (1).
Mielenkiintoista kyllä naiset voittavat miehet, eli väsyvät vähemmän, kun tehdään toistettavia lihassupistuksia samalla suhteellisella intensiteetillä, niin ylä- kuin alaraajoissa (12). Lisäksi naisilla on korkeampi relatiivinen kriittinen intensiteetti, eli he pystyvät pitämään suhteellisesti korkeampaa työtä yllä kuin miehet. Myös naisten korkeampi suhteellinen metabolinen kynnys tarkoittaa, että he väsyvät vähemmän samalla suhteellisella intensiteetillä kuin miehet (13). Tämä tarkoittaa sitä, että naisten pitäisi kenties harjoitella keskiarvollisesti korkeammalla intensiteetillä kuin miesten, kun taas korkein intensiteetti on miehillä suurempi, kuin naisilla. Naisten on myös huomattu väsyvän vähemmän toistuvien juoksuharjoitusten jälkeen (14).
Mistä ero väsymyksessä johtuvat?
Sukupuolten välinen ero aineenvaihdunnassa liittyy lihassolujen suhteelliseen jakaumaan ja sen vaikutuksiin supistumistoiminnassa (14). Miehillä on suurempi glykolyyttinen kapasiteetti kuin naisilla, kun taas naisilla on yleensä suurempi hapenkäyttökapasiteetti koko lihaksessa kuin miehillä (15). Tämän takia esimerkiksi maailmanennätysten erot tempauksessa ovat huomattavasti suuremmat kuin maratonissa.
Myös muut mekanismit vaikuttavat, kuten erot lihasten perfuusiossa ja verenvirtauksessa. Tämä johtuu suonten laajenemisesta ja niiden aktivoinnista, jotka ovat suurempia naisilla kuin miehillä (16). Erot perfuusiossa tai lihassolujen suhteellisessa jakaumassa voivat molemmat johtaa väsymystä aiheuttavien metaboliittien suurempaan ja nopeampaan kertymiseen miehillä suhteessa naisiin väsytystilanteissa, mikä lisää miesten väsyvyyttä verrattuna naisiin.
Naiset loistavat kestävyydessä
Sukupuoliero suorituskyvyssä huippuluokan kestävyysjuoksijoiden välillä maailmanennätyksissä 10 km ja maratonmatkoilla vaihtelee noin 10–12 % välillä. Uinnin maailmanennätyksissä on pienempi sukupuoliero pidemmillä matkoilla (noin 7 % 1500 m matkalla vs. 13 % 50 m vapaauinnissa). Ultramatkan avovesiuinnissa ero on noin 10 % vuoden 2023 maailmanmestaruuskilpailujen 5 km matkalla. Kestävyysurheilussa parhaat miehet suoriutuvat kyllä paremmin kuin parhaat naiset, mutta prosentuaaliset erot ovat pienempiä kuin voima- ja tehourheilussa.
Erot maksimaalisessa hapenotossa johtuvat ainoastaan suuremmasta rasvattomasta massasta?
Tärkeä kestävyyskyvyn ja sukupuolten välisten erojen määräävä tekijä samanikäisten ja samankaltaisesti harjoitelleiden miesten ja naisten välillä on maksimaalinen hapenottokyky. Maksimaalinen aerobinen teho, joka tunnetaan myös maksimaalisena hapenottokykynä, edustaa korkeinta hapenottokykyä ja -käyttöä suorituksen aikana, ja se on keskeinen ihmisen kestävyyskyvyn mittari.
Huippu-urheilijoilla ja aktiivisilla kuntoilijoilla sukupuoliero aerobisessa tehossa vaihtelee noin 20–40 % välillä. Kun se normalisoidaan kehon massaan (ml O2·kg−1·min−1), ero on noin 10–20 % suurempi miehillä kuin naisilla. Mielenkiintoista kyllä, kun tämä normalisoidaan vielä rasvattomaan massaan, niin ero pienenee huomattavasti. Sukupuoliero voi tällöin tippua enää viiteen prosentiin ja jotkut ovat raportoineet lähes merkityksettömiä eroja (17,18). Mitä muita eroja kestävyydessä on sukupuolten välillä?
Tigst Assefa juoksi Berlinissä uuden maratoniin maailman ennätyksen vuonna 2023 aikaan 2:11:53. 70-luvun lopulla maailman ennätys oli 2:30 pinnoilla. Hurja kehitys johtuu siitä, että yhä useampi nainen osallistuu urheilutoimintaan. Tiivistävätkö naiset eroja miehiin kestävyysmatkoilla kun yhä useammalla naisella on mahdollisuus osallistua urheilun pariin?
Naisilla on parempi taloudellisuus juostessa ja uinnissa
Maksimaalinen syke, joka saavutetaan liikunnan aikana, ei kuitenkaan eroa samankaltaisesti harjoitelleiden miesten ja naisten välillä (22). Naisten alempi maksimaalinen hapenottokyky verrattuna miehiin ei johdu siis sykkeistä, vaan pääasiassa siitä, että naisten pienempi sydämen massa ja keuhkojen koko vähentävät kykyä toimittaa hapetettua verta (jossa on vähemmän hemoglobiinia) työskenteleville lihaksille verrattuna samankaltaisesti harjoitelleeseen mieheen.
Mielenkiintoista kyllä useilla eri tasoilla kilpailevilla naisjuoksijoilla on havaittu hieman parempi juoksutalous (alhaisempi hapenkulutus submaksimaalisilla nopeuksilla) kuin samankaltaisesti harjoitelleilla miehillä (23,24).
Uinnissa naiset voittavat taloudellisuudessa miehet huomattavasti (25). Tämä etu vedessä johtuu todennäköisesti naisten korkeammasta kehon rasvaprosentista ja pienemmästä kehon pinta-alasta, mikä vähentää veden vastusta ja siten hapenkulutusta tietyllä nopeudella. Liikkeen taloudellisuus vedessä saattaa selittää miesten edun kaventumisen uinnissa 50 metrin sprinttimatkan ja 1500 metrin pidemmän matkan välillä.
Mitä ollaan opittu näistä tutkimuksista ja käytännön valmennuksen kokemuksesta?
Naiset tarvitsevat noin 10% vähemmän proteiinia kuin miehet (30). Tämä johtuu siitä, että naiset hapettavat vähemmän proteiinia harjoittelun aikana ja kuluttavat vähemmän proteiinia ennen aterioita ja niiden jälkeen.
Naiset kestävät enemmän harjoitusvoluumia. Lihassolutyyppi ei ole ainoa määrittävä tekijä, vaan myös suurempi määrä estrogeeniä vaikuttaa tähän. Estrogeeni on antikatabolinen hormoni, mikä auttaa lihasten korjauksessa ja suojelee lihasvaurioilta. Naisten tulisikin kenties harjoitella lähempänä uupumusta, kuin miesten (33).
Naiset eivät tarvitse yhtä paljon taukoa sarjojen välissä verrattuna miehiin (31).
Kokonaisuudessaan naisten voimaharjoittelussa kannattaa hyödyntää pidempiä sarjoja enemmän kuin miehillä, työskennellä lähempänä uupumusta ja harjoitella hieman tiheämmin kuin miesten kanssa.
Naiset ovat suhteellisesti yhtä voimakkaita kuin miehet, mutta eivät yhtä räjähtäviä (34). Räjähtävästä harjoittelusta voi olla todella haastavaa palautua. Esimerkiksi kovasta juoksuharjoittelusta meni naisilla palautua yli 72 tuntia ja erot miehiin näyttäisivät tulevan harjoituksen jälkeisessä proteiinin muodostamisessa. Juoksujen jälkeen naisten proteiinin tuotto oli selvästi heikompaa kuin miehillä (36), mutta normaalin voimaharjoittelun jälkeen jopa parempaa kuin miehillä (37). Räjähtävä harjoittelu ei siis välttämättä kehitä yhtä tehokkaasti naisia, kuin miehiä, vaan joudutaan keskittymään enemmän suhteelliseen voimantuottoon. Miehillä myös luontainen testosteronin tuotanto auttaa räjähtävän voiman ylläpidossa ja kehittämisessä.
Lähteet
1. Hunter, S. K., & Senefeld, J. W. (2024). Sex differences in human performance. The Journal of physiology, 602(17), 4129–4156. https://doi.org/10.1113/JP284198
2. Hunter, S. K., S Angadi, S., Bhargava, A., Harper, J., Hirschberg, A. L., D Levine, B., L Moreau, K., J Nokoff, N., Stachenfeld, N. S., & Bermon, S. (2023). The biological basis of sex differences in athletic performance: consensus statement for the american college of sports medicine. Medicine and Science in Sports and Exercise, 55(12), 2328–2360.
3. James, J. J., Klevenow, E. A., Atkinson, M. A., Vosters, E. E., Bueckers, E. P., Quinn, M. E., Kindy, S. L., Mason, A. P., Nelson, S. K., Rainwater, K. A. H., Taylor, P. V., Zippel, E. P., & Hunter, S. K. (2023). Underrepresentation of women in exercise science and physiology research is associated with authorship gender. Journal of Applied Physiology (1985), 135(4), 932–942.
4. Morton, R. W., Sato, K., Gallaugher, M. P. B., Oikawa, S. Y., Mcnicholas, P. D., Fujita, S., & Phillips, S. M. (2018). Muscle androgen receptor content but not systemic hormones is associated with resistance training-induced skeletal muscle hypertrophy in healthy, young men. Frontiers in Physiology, 9, 1373.
5. Chapman, M. A., Arif, M., Emanuelsson, E. B., Reitzner, S. M., Lindholm, M. E., Mardinoglu, A., & Sundberg, C. J. (2020). Skeletal muscle transcriptomic comparison between long-term trained and untrained men and women. Cell Reports, 31(12), 107808.
6. Roth, S. M., Ferrell, R. E., Peters, D. G., Metter, E. J., Hurley, B. F., & Rogers, M. A. (2002). Influence of age, sex, and strength training on human muscle gene expression determined by microarray. Physiological Genomics, 10(3), 181–190.
7. Miller, A. E. J., Macdougall, J. D., Tarnopolsky, M. A., & Sale, D. G. (1993). Gender differences in strength and muscle fiber characteristics. European Journal of Applied Physiology and Occupational Physiology, 66(3), 254–262.
8. Frontera, W. R., Suh, D., Krivickas, L. S., Hughes, V. A., Goldstein, R., & Roubenoff, R. (2000). Skeletal muscle fiber quality in older men and women. American Journal of Physiology-Cell Physiology, 279(3), C611–C618.
9. Grosicki, G. J., Zepeda, C. S., & Sundberg, C. W. (2022). Single muscle fibre contractile function with ageing. The Journal of Physiology, 600(23), 5005–5026.
10. Esbjörnsson-Liljedahl, M., Sundberg, C. J., Norman, B., & Jansson, E. (1999). Metabolic response in type I and type II muscle fibers during a 30-s cycle sprint in men and women. Journal of Applied Physiology, 87(4), 1326–1332.
11. Staron, R. S., Hagerman, F. C., Hikida, R. S., Murray, T. F., Hostler, D. P., Crill, M. T., Ragg, K. E., & Toma, K. (2000). Fiber type composition of the vastus lateralis muscle of young men and women. Journal of Histochemistry and Cytochemistry, 48(5), 623–629.
12. Ansdell, P., Thomas, K., Hicks, K. M., Hunter, S. K., Howatson, G., & Goodall, S. (2020). Physiological sex differences affect the integrative response to exercise: Acute and chronic implications. Experimental Physiology, 105(12), 2007–2021.
13. Ansdell, P., Brownstein, C. G., Škarabot, J., Hicks, K. M., Howatson, G., Thomas, K., Hunter, S. K., & Goodall, S. (2019). Sex differences in fatigability and recovery relative to the intensity-duration relationship. The Journal of Physiology, 597(23), 5577–5595.
14. Billaut, F., & Bishop, D. (2009). Muscle fatigue in males and females during multiple-sprint exercise. Sports Medicine (Auckland, N.Z.), 39(4), 257–278.
15. Esbjörnsson, M., Sylvén, C., Holm, I., & Jansson, E. (1993). Fast twitch fibres may predict anaerobic performance in both females and males. International Journal of Sports Medicine, 14(05), 257–263.
16. Hunter, S. K., Butler, J. E., Todd, G., Gandevia, S. C., & Taylor, J. L. (2006). Supraspinal fatigue does not explain the sex difference in muscle fatigue of maximal contractions. Journal of Applied Physiology (1985), 101(4), 1036–1044.
17. Bunc, V., & Heller, J. (1989). Energy cost of running in similarly trained men and women. European Journal of Applied Physiology and Occupational Physiology, 59(3), 178–183.
18. Cureton, K., Bishop, P., Hutchinson, P., Newland, H., Vickery, S., & Zwiren, L. (1986). Sex difference in maximal oxygen uptake. Effect of equating haemoglobin concentration. European Journal of Applied Physiology and Occupational Physiology, 54(6), 656–660.
19. Dominelli, P. B., Molgat-Seon, Y., & Sheel, A. W. (2019). Sex differences in the pulmonary system influence the integrative response to exercise. Exercise and Sport Sciences Reviews, 47(3), 142–150.
20. Ripoll, J. G., Guo, W., Andersen, K. J., Baker, S. E., Wiggins, C. C., Shepherd, J. R. A., Carter, R. E., Welch, B. T., Joyner, M. J., & Dominelli, P. B. (2020). Sex differences in paediatric airway anatomy. Experimental Physiology, 105(4), 721–731.
21. Sheel, A. W., Guenette, J. A., Yuan, R., Holy, L., Mayo, J. R., Mcwilliams, A. M., Lam, S., & Coxson, H. O. (2009). Evidence for dysanapsis using computed tomographic imaging of the airways in older ex-smokers. Journal of Applied Physiology (1985), 107(5), 1622–1628.
22. Bassareo, P. (2020). Gender differences in hemodynamic regulation and cardiovascular adaptations to dynamic exercise. Current Cardiology Reviews, 16(1), 65–72.
23. Støa, E. M., Helgerud, J., Rønnestad, B. R., Hansen, J., Ellefsen, S., & Støren, Ø. (2020). Factors influencing running velocity at lactate threshold in male and female runners at different levels of performance. Frontiers in Physiology, 11, 585267.
24. V Mendonca, G., Matos, P., & Correia, J. M. (2020). Running economy in recreational male and female runners with similar levels of cardiovascular fitness. Journal of Applied Physiology (1985), 129(3), 508–515.
25. Pendergast, D. R., Di Prampero, P. E., Craig, A. B., Jr., Wilson, D. R., & Rennie, D. W. (1977). Quantitative analysis of the front crawl in men and women. Journal of Applied Physiology Respiratory, Environmental Exercise Physiology, 43, 475–479.
26. Cano, A., Ventura, L., Martinez, G., Cugusi, L., Caria, M., Deriu, F., & Manca, A. (2022). Analysis of sex-based differences in energy substrate utilization during moderate-intensity aerobic exercise. European Journal of Applied Physiology, 122(1), 29–70.
27. Horton, T. J., Pagliassotti, M. J., Hobbs, K., & Hill, J. O. (1998). Fuel metabolism in men and women during and after long-duration exercise. Journal of Applied Physiology (1985), 85(5), 1823–1832.
28. Chapman, M. A., Arif, M., Emanuelsson, E. B., Reitzner, S. M., Lindholm, M. E., Mardinoglu, A., & Sundberg, C. J. (2020). Skeletal muscle transcriptomic comparison between long-term trained and untrained men and women. Cell Reports, 31(12), 107808.
29. Joseph et al. (2014) Where are all the female participants in Sports and Exercise Medicine research?, European Journal of Sport Science, 14:8, 847-851.
30. Rand, W. M., Pellett, P. L., & Young, V. R. (2003). Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. The American journal of clinical nutrition, 77(1), 109–127. https://doi.org/10.1093/ajcn/77.1.109
31. Fulco, C. S., Rock, P. B., Muza, S. R., Lammi, E., Cymerman, A., Butterfield, G., Moore, L. G., Braun, B., & Lewis, S. F. (1999). Slower fatigue and faster recovery of the adductor pollicis muscle in women matched for strength with men. Acta physiologica Scandinavica, 167(3), 233–239. https://doi.org/10.1046/j.1365-201x.1999.00613.x
32. Maughan, R. J., Harmon, M., Leiper, J. B., Sale, D., & Delman, A. (1986). Endurance capacity of untrained males and females in isometric and dynamic muscular contractions. European journal of applied physiology and occupational physiology, 55(4), 395–400. https://doi.org/10.1007/BF00422739
33. Rissanen, J., Walker, S., Pareja-Blanco, F., & Häkkinen, K. (2022). Velocity-based resistance training: do women need greater velocity loss to maximize adaptations?. European journal of applied physiology, 122(5), 1269–1280. https://doi.org/10.1007/s00421-022-04925-3
34. Häkkinen K. (1991). Force production characteristics of leg extensor, trunk flexor and extensor muscles in male and female basketball players. The Journal of sports medicine and physical fitness, 31(3), 325–331.
35. Keane, Karen M.; Salicki, Rebecca; Goodall, Stuart; Thomas, Kevin; Howatson, Glyn. Muscle Damage Response in Female Collegiate Athletes After Repeated Sprint Activity. Journal of Strength and Conditioning Research 29(10):p 2802-2807, October 2015. | DOI: 10.1519/JSC.0000000000000961
36. Scalzo, R. L., Peltonen, G. L., Binns, S. E., Shankaran, M., Giordano, G. R., Hartley, D. A., Klochak, A. L., Lonac, M. C., Paris, H. L., Szallar, S. E., Wood, L. M., Peelor, F. F., 3rd, Holmes, W. E., Hellerstein, M. K., Bell, C., Hamilton, K. L., & Miller, B. F. (2014). Greater muscle protein synthesis and mitochondrial biogenesis in males compared with females during sprint interval training. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 28(6), 2705–2714. https://doi.org/10.1096/fj.13-246595
37. Henderson GC, Dhatariya K, Ford GC, Klaus KA, Basu R, Rizza RA, Jensen MD, Khosla S, O’Brien P, Nair KS. Higher muscle protein synthesis in women than men across the lifespan, and failure of androgen administration to amend age-related decrements. FASEB J. 2009 Feb;23(2):631-41. doi: 10.1096/fj.08-117200. Epub 2008 Sep 30. PMID: 18827019; PMCID: PMC2630787.
38. Judge, L. W., & Burke, J. R. (2010). The effect of recovery time on strength performance following a high-intensity bench press workout in males and females. International journal of sports physiology and performance, 5(2), 184–196. https://doi.org/10.1123/ijspp.5.2.184
39. Refalo, M.C.,Nuckols, G., Galpin, A.J.,Gallagher, I.J., Hamilton, D.L., & Fyfe, J.J (2024). Sex Differences in Absolute and Relative Changes in Muscle Size following Resistance Training in Healthy Adults: A Systematic Review with Bayesian Meta-Analysis. SportRχiv
Haluatko ostaa tarkimmat mittausvälineet mitä markkinoilla on hyppykorkeuden tai vertikaalisten voimien mittaamiseksi? Olet tullut oikeaan paikkaan. Tämä opas auttaa sinua hankkimaan juuri sopivat voimalevyt käyttötarkoitukseesi.
Mitkä ihmeen voimalevyt?
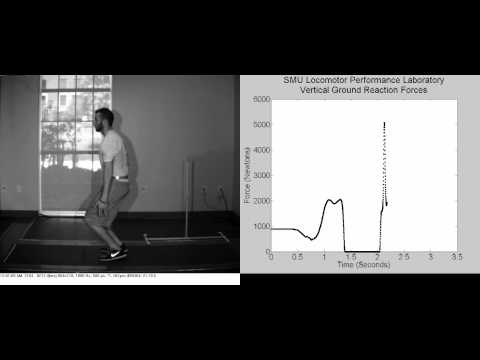
Voimalevyillä saa tarkasti mitattua esimerkiksi hyppykorkeuden tai kuinka nopeasti maksimivoimatestissä tuotetaan voimaa. Voihan hyppykorkeuden mitata optisilla laitteilla tai kännykkäsovelluksellakin, mutta näissä lasketaan hyppykorkeus lentoajasta, kun taas voimalevyt laskevat hyppykorkeuden tuotetusta voimasta levyjä vastaan. Esimerkiksi Optojump - mittausjärjestelmän ja voimalevyjen välillä on selkeä ero (-1.06 cm; p < 0.001), vaikkakin Optojump on todettu muuten suhteellisen luotettavaksi mittausjärjestelmäksi (Glatthorn et al., 2011).
Voimalevyjen toiminta on suhteellisen yksinkertaista. Esimerkiksi hyppykorkeuden mittaamiseksi asetetaan voimalevy hyppijän alle, ja kun hyppijä ponnistaa, niin paine ja paino kohdistuvat levylle ja muuntuvat sähköisiksi signaaleiksi. Nämä signaalit voidaan sitten tallentaa ja analysoida ja niiden avulla voidaan määrittää hyppykorkeus. Tämä perustuu ajatukseen, että hyppykorkeus on suoraan verrannollinen levyn pinnalle kohdistuvaan painoon ja siitä aiheutuvaan sähköiseen signaaliin.
Kuva 1: Kistlerin kannettavat voimalevyt
Voimalevyjen avulla määritetty hyppykorkeus on tarkka ja luotettava, ja ne ovat olleet tärkeitä välineitä urheilun ja tieteellisen tutkimuksen saralla jo kohta 40 vuotta. Niitä käytetään laajasti esimerkiksi urheilu- ja liikuntatutkimuksessa. Ennen vain yliopistot ja isot seurat pystyivät haaveilemaan niiden ostamisesta, mutta tekniikan kehittyessä hinnat ovat tulleet alas nopeasti, samaan tapaan kuin tangon liikenopeusmittareiden kanssa on käynyt.
Kuva 2: Optiset järjestelmät, kuten Optojump ovat hyvin suosittuja laitteita hyppykorkeuden määrittelemiseksi. Ne laskevat hyppykorkeuden lentoajasta ja alastulotekniikka vaikuttaa merkittävästi hyppytulokseen, toisin kuin voimalevyillä. Isommilla ulkoisilla kuormilla tehtävät hypyt ovat hankalia kontrolloida optisilla järjestelmillä.
Mitä voimalevyillä voi mitata?
Voimalevyt mahdollistavat urheilijoiden maahan kohdistamien voimien mittaamista liikkumisen tai harjoittelun aikana. Voimalevyt ovat yksinkertaisen näköisiä neliöitä tai suorakaiteita, mutta ne voidaan upottaa myös maahan tasaisesti (kuva 4). Voimalevyjä käytetään urheilijoiden maan reaktiovoiman analysointiin kävellessä, juostessa, hyppiessä tai muissa fyysisissä testeissä.
Voimalevyillä voidaan mitata muun muassa:
Nopeutta (m/s)
Tehoa (watit)
Siirtymää (cm)
Ajallisia parametreja (s)
Vasen/oikea asymmetria (vain tuplalevyissä)
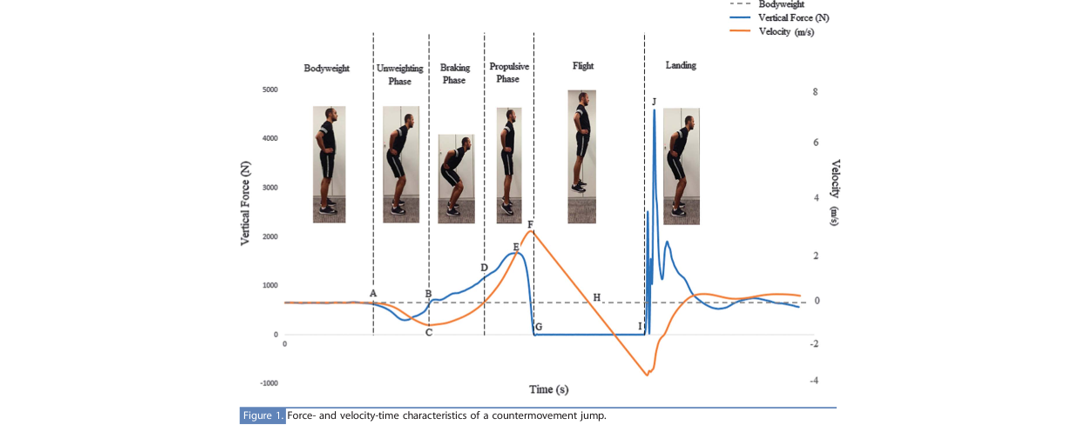
Voimalevyillä tehtäessä hyppytestejä voidaan hyppyvaihe jakaa eri osiin tehokkaasti toisin kuin optisilla järjestelmillä. Tällöin voidaan analysoida hypyn eri vaiheita ja osoittaa tehokkaammin harjoitusärsykkeitä juuri halutuille hypyn vaiheille. Optisilla järjestelmillä saat vain hyppykorkeuden tietoosi.
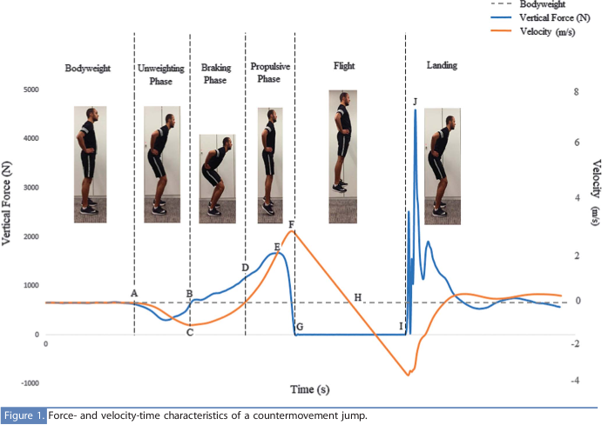
Kuva 3: Hypyn eri osavaiheet esitettynä kuvassa vertikaalisen voimakäyrän ja liikenopeuden kanssa (Chavda et al., 2017).
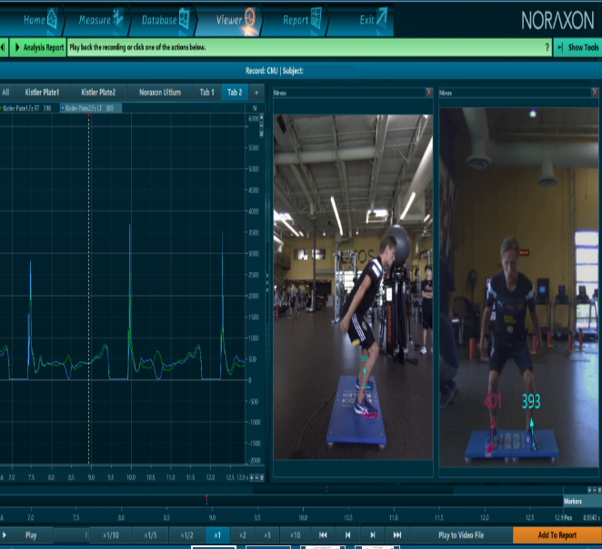
Hyppääminen ja datan kerääminen näyttää Hawkinin levyillä tehtyinä tältä:
Levyjä voi käyttää myös yläkropan nopeusvoimaominaisuuksien mittaamiseen. Tässä esimerkiksi Hawkinin levyt ja punnerruhyppy.
Voimalevyjen avulla voidaan määrittää lukuisia eri indeksejä, joista mielenkiintoisimmat ovat mielestäni:
Dynaaminen Voimaindeksi (DSI) - ero urheilijan maksimaalisessa ja räjähtävässä voimassa.
Voimalevyillä päästään siis pintaa syvemmälle verrattuna pelkkään hyppykorkeuteen. Levyt mahdollistavat yksilöllisemmän analyysin tekemisen eri testeissä.
Voimalevyjen tekniikka ja tarkkuus?
Voimalevyjen suositeltu näytetaajuus on 1000 Hz, joten voimalevyt mittaavat siihen kohdistuneita voimia 1 millisekunnin välein. Jos tuotteessa on alle tuhannen hertsin mittaustarkkuus, niin pyytäisin aina selitystä tälle tuotteen valmistajalta. Alhaisella mittaustarkkuudella voi osa nopeiden liikkeiden huippuarvoista mennä ohi.
Voimalevyt mittaavat myös voimia, vaikkei mitään ole niiden päällä. Tätä kutsutaan kohinaksi, ja se on läsnä kaikissa voimalevyissä. Kohina halutaan pitää tietenkin mahdollisimman matalana, jotta mittausten tarkkuus ja sensitiivisyys paranee. On siis tärkeää nollata voimalevyt joka kerta kuin urheilija astuu levyille. Onneksi lähes kaikkien nykyaikaisten voimalevyjen ohjelmistot nollaavat kohinan automaattisesti. Voimalevyjen tulee sijaita myös tasaisella ja tukevalla pinnalla mittauksen aikana.
Mitattavan urheilijan paino tulee mitata joka kerta. Ilman painoa ei voida laskea mitään ja silloin voimalevyt ovat yhtä tehokkaita kuin normaalit hyppymatot. Lähes kaikki nykyaikaiset voimalevyt mittaavat mitattavan painon automaattisesti.
Kuva 4: Voimalevyjä voi upottaa myös radan pintaan tai vaikka uimarin lähtötelineeseen. Esimerkiksi Japanissa on 100m juoksurata pelkkää voimalevyä. Tälläisellä saa hyvin tarkasti tehtyä tieteellistä tutkimusta pikajuoksusta. Valistunut arvioni on tämmöisen kokonaisuuden hinnaksi reippaasti yli miljoona euroa. Yksityisyrittäjälle riittää pienempikin investointi.
Tarkkuus?
Voimalevyt ovat todella tarkkoja ja esimerkiksi Hawking Dynamics ilmoittaa itse tarkkuudeksi 0,1N (newton) ja +-0.25N täsmällisyydeksi. Tarkkuus viittaa siihen miten lähellä mittaustulos on todellista lukua ja täsmällisyys mittaa kuinka lähellä eri mittauskerrat ovat toisistaan. Bertecin levyissä vaihteluväli näyttäisi olevan +3 ja -2N välillä keskiarvon ollessa 1N. Tämä näyttäisi olevan valmistajan ilmoittama tarkkuus lähes kaikissa levyissä. 1N virhe on hyvin mitätön esimerkiksi 350N paineella (350N on tyypillinen luku kun 70 kg ihminen seisoo tasaisesti levyjen päällä). Ja mitatessa hyppyjä tai muita voimatestejä, jossa voidaan pyöriä esimerkiksi 2kN - 4kN välillä. Tällöin 1N on lähes merkityksetön virhemarginaali. Kannattaa huomioida, että valmistajien ilmoittamat virhemarginaalit vaihtelevat huomattavasti. Esimerkiksi ilmoitettu virhemarginaali voi vaihdella 0,075 N - 0,2N välillä. Käytännön työssä erot ovat mitättömiä, koska 0,1N vastaa pyöristettynä noin 0.0101971621 kilogrammaa.
Mittauskertojen välinen ero esimerkiksi plyometrisissa punnerruksissa näyttäisi olevan erittäin tarkka. Punnerruksissa mitattiin voimalevyjen variaatiokertoimeksi (coefficient of variation, CV) CV = 2.3%–11%. Luku suhteuttaa keskihajonnan aineiston keskiarvoon ja erittäin karkeasti CV < 15% on erittäin luotettava ja CV > 30 kertoo siitä, että mittauskertojen ero on liian suuri, jotta siihen kannataisi luottaa. Merijalkaväen sotilailla mitattiin myös erittäin tarkkaa vaihtelua (CV = 4.8%) (Hogarth et al., 2013; Hrysomalis & Kidgell, 2001; Koch et al., 2012).
Mitä muuta kannattaa ottaa huomioon?
Bilateraalinen vai yksittäinen voimalevy?
Jos haluat mitata raajojen välistä eroa tai asymetriaa, niin hanki bilateraaliset levyt.
Tarvitsetko 3D analyysiä?
Et tarvitse, ellet ole tutkija. Useimmat voimalevyt mittaavat vain vertikaalisia voimia. Jos haluat mitata voimia horisontaalisesti tai lateraalisesti varaudu investoimaan huomattavasti enemmän rahaa. Datan tulkinta on myös tällöin huomattavasti hankalampaa ja vaatii enemmän aikaa. Suosittelen tätä vain tutkijoille.
Kannettavuus?
Paljonko levyt painavat? Kannatko niitä paljon paikasta toiseen vai seisovatko levyt yhdessä paikassa jatkuvasti? Paino on merkittävä asia valmentajalle, jonka toimipiste liikkuu urheilijoiden mukana.
Sovellus
Useimmissa voimalevyjärjestelmissä on nykyään ohjelmisto, jonka avulla voit tulkita raakadataa. Tämä ei kuitenkaan tarkoita, että kaikki ohjelmistot ovat samankaltaisia. Haluatko tiedot kätevästi suoraan puhelimeen vai haluatko tulkita dataa tietokoneen näytöltä. Käyttötarkoitus vaikuttaa huomattavasti tähän valintaan. Lisäksi toinen iso asia on, että saako raakadataa ladattua itselleen esimerkiksi exceliin?
Kuinka paljon tukea tarvitset?
Tarvitsetko paljon tukea käytön aloittamiseen tai käytössä? Ota huomioon onko voimaleyjä tarjoavalla yrityksellä oma tukitiimi? Ovatko nämä tukijäsenet koulutettuja järjestelmästään, ja he ymmärtävät myös kuinka urheilijoita valmennetaan?
Kuva 5: Bertecin voimalevyistä ja ohjelmistosta kuva.
Markkinoiden parhaat vaihtoehdot?
Onko sensorilla väliä?
Vastusvenymäliuska on kaikkein yleisin sensorityyppi. Pietsosähköiset voima-anturit ovat selvästi kalliimpia. Näissä voima kohdistetaan pietsosähköiseen kristalliin, mikä aiheuttaa varaussiirtymän molekyylitasolla ja hilarakenteen sisällä. Tämä sähkövaraus vangitaan kiteen pinnalle ja muunnetaan jännitesignaaliksi ns. varausvahvistimen avulla. Tämän menetelmän edut ovat äärimmäisen nopeiden tai korkeataajuisten mittaustapahtumien tallentamisessa. Toisaalta sähkövaraus on tässä menetelmässä epävakaa ja jos täydellistä eristystä ei ole, lataus katoaa ajan myötä. Tämä tekee pitkäaikaisesta vakaasta mittauksesta vaikeaa, varsinkin silloin, jos halutaan mitata pienehköjä voimia. Lisäksi lämpötilavaihtelut vaikuttavat enemmän pietsosähköiseen anturiin. Fysiikkavalmennus.fi suosittelee siis vastusvenymäliuskoja kaikille muille paitsi yliopistotason tutkijoille.
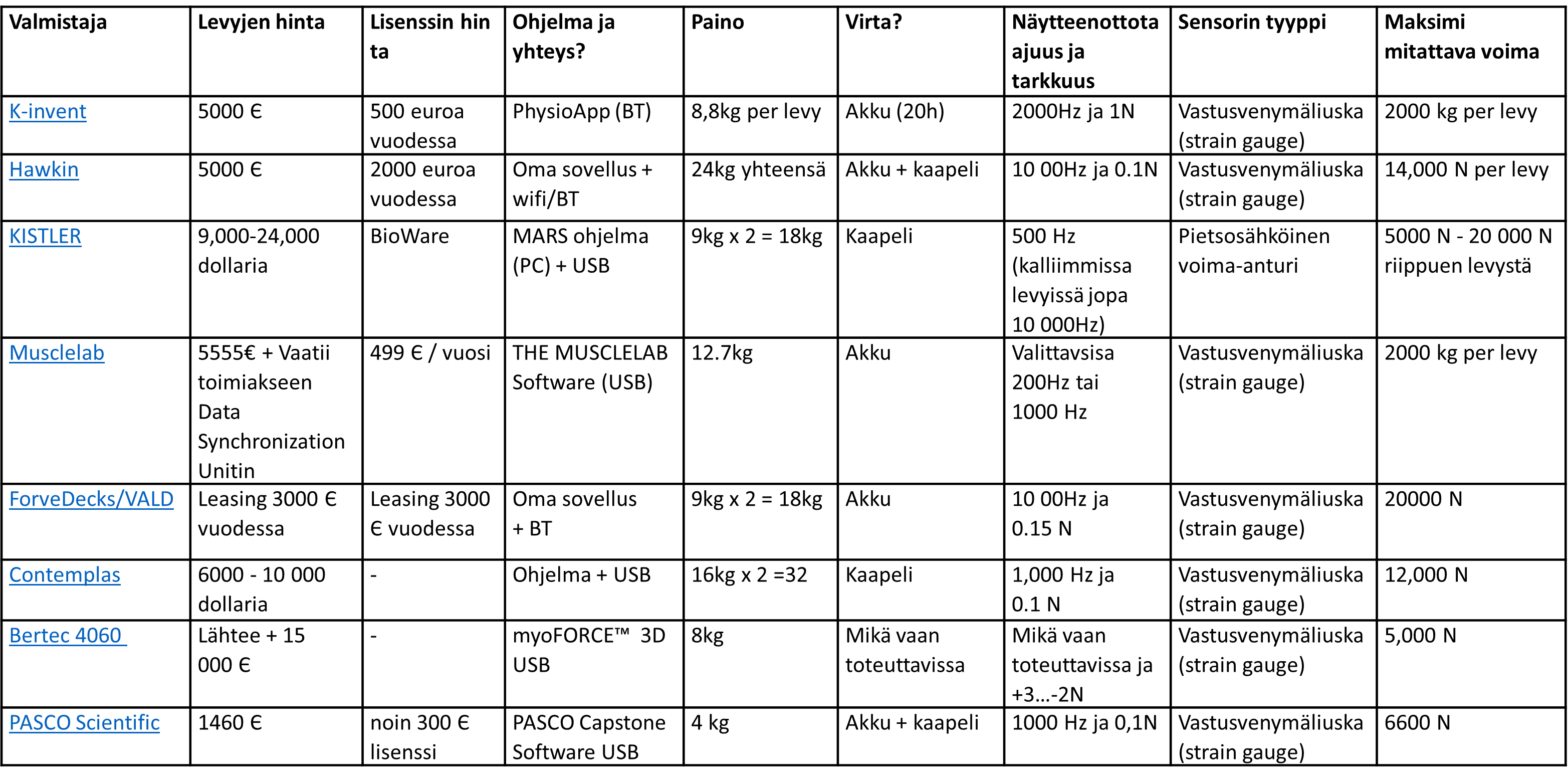
Alla olevassa taulukossa on esitelty fysiikkavalmennus.fi valitsemia voimalevyjä ja niiden eri ominaisuuksia (taulukkoa voi mobiilin selainversiolla zoomata). Huomioi, että laitteet ja ohjelmistot päivittyvät kokoajan, joten tämä lista ei välttämättä ole täysin ajantasainen.
Suositukset ja käyttökokemukset
Musclelabilla pystyy yhdistämään Data Synchronization Unitilla esimerkiksi EMG:n ja voimalevyjen datan. Sillä pystyy yhdistämään myös muut Musclelabin laitteet, kuten tangon liikenopeusmittarin. Jos aiot rakentaa kunnon mittaustehtaan, niin suosittelen vahvasti Musclelabin laitekokonaisuuksia.
Vald voimalevyt ovat tulleet ryminällä markkinoilla ja yritys on laajentanut nopeasti myös Suomeen. Useat valmentajat ovat ostaneet nämä voimalevyt ja käyttökokemus on kuulemani mukaan kaikilla ollut positiivinen. Vahva suositus näille, jos levyt ja lisenssin maksaa joku muu kuin sinä. Esimerkiksi seuralle tai urheiluopistolle loistava valinta. Yksityisyrittäjällä 3000 euron vuosittainen leasing maksu voi olla liian suolainen pala.
Itse ostin K-inventin levyt, koska levyt sai kerralla itselleen ja lisenssi ei ole törkeän kallis. Esimerkiksi verrattuna Hawkinin lisenssiin (2000 euroa vuodessa) K-inventin lisenssit ovat halvat. Olen itse tykännyt kovasti levyistä. Ohjelmisto on pääasiassa suuniteltu fysioterapeuteille, kuten nimikin viittaa: PhysioApp. Mutta valmistaja vakuutti, että ovat rakentamassa PerformanceApp - ohjelmistoa myös fysiikkavalmentajille. Nykyinen ohjelmisto on kömpelö isompien joukkueiden testaamiseen, mutta soveltuu hyvin yksittäisten urheilijoiden mittaamiseen.
Kistlerin ja Bertecin voimalevyt ovat muokattavissa mihin ympäristöön vain ja yritykset toteuttavat mittatilaustuotteena upotettuja ja perinteisiä levyjä. Kummankin yrityksen levyt ovat huomattavasti kalliimpia kuin minkään muun yrityksen. Yliopistot käyttävät lähes poikkeuksetta Kistleriä. Kistleri käyttää pietsosähköisiä voima-antureita levyissään, joten hintakin on huomattavasti kalliimpi.
Musta hevonen
Oletko kätevä käsistäsi ja ymmärrät ohjelmoinnin päälle? Osta ehdottomasti chronojumpilta 250 euroa maksavat voimasensorit (2-4 riittää) ja rakenna levy itse. Ohjelmisto antaa ihan hyvin raakaa dataa, mutta vaatii ymmärrystä ja osaamista käytöstä, joten hinta-laatu suhteeltaan tämä on osaavalle yksilölle ehdottomasti paras valinta.
Fysiikkavalmennus.fi suosittelee
Seurat, opistot, koulut ja muut isommat toimijat:VALD - voimalevyt
Fysiikkavalmentajat: Musclelab tai K-Invent kakkosvaihtoehtona
Tutkijat ja yliopistot: Bertec tai Kistler
Tee-se-itse: Chronojump
Lähteet
Glatthorn JF, Gouge S, Nussbaumer S, Stauffacher S, Impellizzeri FM, Maffiuletti NA. Validity and reliability of Optojump photoelectric cells for estimating vertical jump height. J Strength Cond Res. 2011;25(2):556-560. doi:10.1519/JSC.0b013e3181ccb18d
Chavda, S., Bromley, T., Jarvis, P., Williams, S., Bishop, C., Turner, A., Lake, J.P., & Mundy, P.D. (2017). Force-Time Characteristics of the Countermovement Jump: Analyzing the Curve in Excel. Strength and Conditioning Journal, 40, 67–77.
Hogarth L, Deakin G, Sinclair W. Are plyometric push-ups a reliable power assessment tool? J Aust Strength Cond. 2013;21:67–69.
Hrysomallis C, Kidgell D. Effect of heavy dynamic resistive exercise on acute upper-body power. J Strength Cond Res. 2001;15(4):426–430. PubMed ID: 11726252
Parry, G. N., Herrington, L. C., & Horsley, I. G. (2020). The Test–Retest Reliability of Force Plate–Derived Parameters of the Countermovement Push-Up as a Power Assessment Tool, Journal of Sport Rehabilitation, 29(3), 381-383. Retrieved Feb 8, 2023, from https://journals.humankinetics.com/view/journals/jsr/29/3/article-p381.xml
Oletko tehnyt harjoitusohjelman ja huomannut ohjelman jälkeen, ettet kehittynytkään tai urheilijasi ei saavuttanutkaan haluttua kehitystä? Kaikki yksilöt kehittyvät eri tahtiin ja tiedämme, että yksilöiden välillä on paljon eroja kehittymistahdissa. Jotkut yksilöt eivät vastaa tietynlaiseen harjoitteluun ollenkaan, kun toiset taas kehittyvät vimmatusti. Lihasmassaharjoittelun yhteydessä käytetään kansankielisesti termistöä hardgaineri, kun kehitystä ei tule. Mitä tämmöisissä tapauksissa kannattaa tehdä, jotta kehitystä saadaan jatkossa aikaan vai onko hardgainer tuomittu loppuelämäksi olemaan kehittymättä?
Mitkä kaikki vaikuttavat kehittymiseen voimaharjoittelussa?
Vaikka voimaharjoittelu onkin erinomainen tapa kehittää lihasvoimaa ja -kestävyyttä, se ei kuitenkaan ole aina yhtä tehokas kaikille ihmisille. Kehittymiseen vaikuttavat monet eri tekijät, kuten ikä, sukupuoli, perimä, elämäntavat, harjoitustausta ja mahdolliset sairaudet tai vammat.
Tämän vuoksi onkin tärkeää ymmärtää ja suunnitella voimaharjoittelua yksilöllisten tarpeiden mukaan. Yksilöllisyyden huomioiminen voimaharjoittelussa auttaa saavuttamaan parempia tuloksia ja välttämään mahdollisia vammoja tai ylirasitusta.
Erittäin tärkeä tekijä kehittymisessä on harjoitustausta. Esimerkiksi vasta-alkaja kehittyy huomattavia määriä alussa lihasmassan ja voiman osalta, kun taas kokeneemmalla harjoittelijalla pienenkin kehityksen saavuttaminen on haastavampaa (Rhea et al., 2003).
Lihaskasvusta ja kaikesta harjoittelusta puhuttaessa ei voi olla puhumatta ravitsemuksesta ja proteiinin merkityksestä. Yksilöt, jotka saavat riittävästi energiaa ja proteiinia tukemaan heidän harjoitteluaan, kehittyvät huomattavasti enemmän kuin ne, jotka eivät saa tarvitsemaansa ravintoa (Phillips and Van Loon, 2011). Voiko syyttää vain geenejään vai onko taustalla jotain muuta?
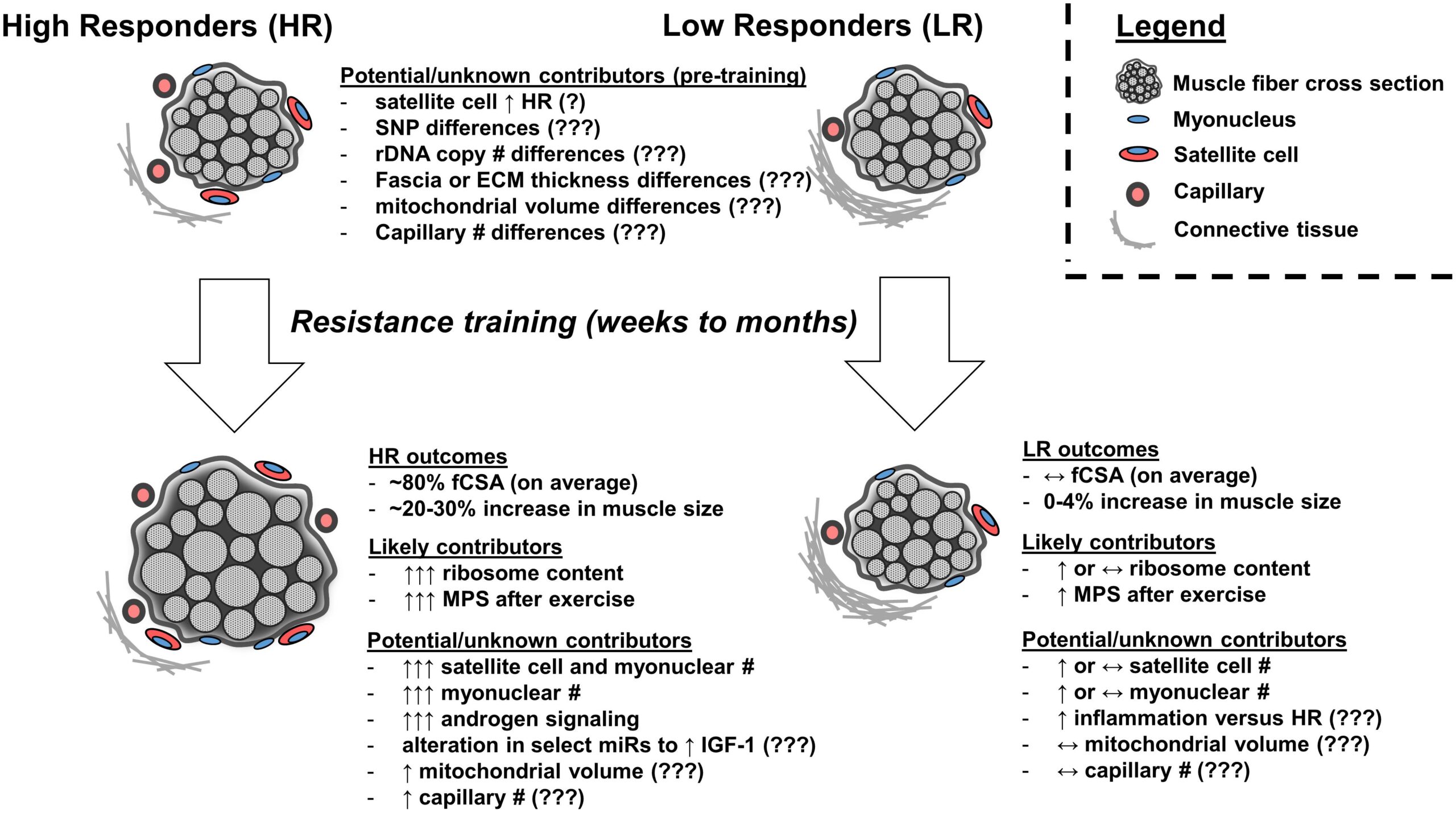
Kuva 1: Nopeasti kehittyvillä yksilöillä näyttäisi olevan suurempi ribosomipitoisuus ja lihasproteiinisynteesi harjoitusten jälkeen verrattuna heikosti kehittyviin yksilöihin. Lisäksi hypoteettisesti hyvin kehittyvillä on enemmän satelliittisoluja, tumia, androgeenisignalointia, mitokondrioiden volyymia ja hiusverisuonistoa. Kuvan lähde Roberts et al., 2018.
Elämäntavat tärkeämpiä kuin geenit?
Genetiikka on eittämättä tärkeässä roolissa yksilön kehityksessä. Esimerkiksi ACTN3 geenin on näytetty olevan yhteydessä eroihin lihasvoiman ja tehontuoton kehittymisessä harjoittelussa (Jones et al., 2016; Eynon et al., 2013). Toisaaltaidenttisillä kaksosilla huomattiin, että elintavoilla on valtava merkitys. Identtiset kaksoset ovat perimältään ja genotyypiltään täysin samanlaiset. Perimän ollessa identtinen, vain olosuhteet vaikuttavat kehitykseen ja näin kaksostutkimukset toimivat esimerkkinä olosuhteiden vaikutuksesta yksilön kehitykseen. 52 vuotiaista kaksosista toinen oli harjoitellut kestävyystyyppisesti yli 30 vuotta ja toinen kaksosista ei ollut liikkunut juuri lainkaan. Harjoitelleella kaksosella oli pienempi kehonmassa, rasvamassa, leposyke, verenpaine, kolesteroli, plasman triglyseridi- ja glukoosipitoisuus. Lisäksi hänellä oli korkeampi anaerobinen ja aerobinen kestävyyskyky. Toisaalta hänellä oli vähemmän lihasta ja voimaa verrattuna ei-harjoitelleeseen kaksoseen. Hänellä oli enemmän hitaita lihassoluja ja vähemmän nopeita lihassoluja sekä korkeampi AMPK proteiinin esiintyvyys (kestävyyssignalointiin liitetty väylä). Yhteensä kestävyysharjoitelleella kaksosella oli 55% enemmän hitaita lihassoluja ja jopa 12,4 ml/kg/min korkeampi VO2max ja 8,6% matalampi rasvaprosentti.
Tutkimuksessa korostettiin elintapojen ja harjoittelun voimakasta vaikutusta koko kehoon. Geenit määrittävät tietyn taustan harjoittelulle, mutta riittävällä harjoittelulla ja elintavoilla voidaan vaikuttaa erittäin tehokkaasti haluttuihin adaptaatioihin. Myös muissa kaksoistutkimuksissa on huomattu (Marsh et al., 2020), etteivät geenit välttämättä ole ihan niin tärkeässä roolissa harjoitusadaptaatiossa kuin aikaisemmin on arvioitu eri poikittaistutkimuksissa.
Toisaalta Davidsenin ja kumppanien tutkimus osoitti, että mikroiRNA-muutokset alhaisen vasteen ihmisillä voivat estää kasvua ja uudistumista edistävien geenien “aktivoitumisen”. Heidän mukaansa lihaskasvu liitty mikroRNA pitoisuuksien muutoksiin ja heidän analyysinsa viittaa siihen, että mikroRNA:lla voi olla rooli voimaharjoittelun aiheuttamissa muutoksissa ja siinä kehittyykö ihminen vai ei. Näyttäisikin siltä, että niin geeneillä kuin olosuhteilla on merkitystä.
Kuva 2: Intohimon lisäksi myös geenit ja olosuhteet vaikuttavat harjoitusadaptaatioiden määrään.
Mitä tehdä jos kehitystä ei tule - lisää harjoituskuormaa?
Harjoituskuormaa, eli volyymia, on pidetty tärkeimpänä tekijänä morfologisia muutoksia varten (Figueiredo et al., 2018). Mattocks et al. (2017) huomasivat, että koehenkilöt, jotka suorittivat hyvin korkean intensiteetin voimaharjoituksia saavuttivat saman voimatasojen kasvamisen kuin ne, jotka harjoittelivat suuremmalla volyymilla. Kuitenkin vain korkeamman volyymin harjoitteluryhmä saavutti lihasmassan kasvua.
Molekyylitason näkökulmasta suurempi harjoitusvolyymi vaikuttaa myofibrillaarisen proteiinisynteesin anabolisen signalointiin positiivisesti. Suurempi volyymi on myös tärkeä p70S6k- ja S6-fosforylaation kannalta voimaharjoittelun jälkeen (Burd ym., 2010a, b; Terzis ym., 2010). p70S6k- ja S6-fosforylaatio säätelevät muun muassa solujen kasvua vaikuttamalla proteiinisynteesin komponentteihin.
Kuinka paljon volyymia on tarpeeksi?
Meta-analyysi ikääntyvien ihmisten lihasmassaharjoittelusta osoitti, että merkittävämmät muutokset liittyivät erityisesti harjoitusohjelmiin, jossa tehtiin enemmän sarjoja koko harjoitussession aikana (Peterson ym., 2011). Useissa tutkimuksissa on huomattu useamman sarjan ylivoimaisuus verrattuna yhteen sarjaan sekä nuorilla että vanhemmilla yksilöillä (Kramer, 1997; Radaelli ym., 2014a,b). Myös Kriegerin (2010) tekemässä meta-analyysissä huomattiin, että usean sarjan suorittaminen on todella tehokkaampaa kuin yhden sarjan suorittaminen. Toisaalta samassa analyysissä huomattiiin, että neljän tai kuuden sarjan suorittaminen ei ole parempaa kuin kahden tai kolmen sarjan suorittaminen. Eli harjoitusvolyymin lisäyksellä voisi olla teoreettinen maksimi yksilön kohdalla. Lisäksi vaikuttaa siltä, ettei supistustapa ole yhtä tärkeä kuin volyymi, jos halutaan saavuttaa molekyylitason muutoksia harjoitussession jälkeen (Garma ym., 2007).
Tietääksemme vain yksi tutkimus on yrittänyt vastata suoraan siihen, voidaanko harjoitusvolyymilla vaikuttaa yksilöllisiin eroihin. Nunes ym. (2021b) huomasivat, että heikosti kehittyvät ihmiset eivät kehittyneet, vaikka harjoitusvolyymia nostettiin. Toisaalta tämä oli retrospektiivinen data-analyysi, eikä suora interventiotutkimus. Scarpelli et al. (2020) näyttivät myös, että ei-optimi harjoitusvolyymi haittaa lihaskasvua nuorilla harjoitelleilla yksilöillä. Toisaalta Montero & Lunby (2017) huomasivat mielenkintoisessa tutkimuksessaan, että kaikki heikosti harjoitteluun vastanneet yksilöt muuttuivat positiivisesti kehittyneiksi yksilöiksi kun harjoittelua lisättiin runsaasti kahdelle seuraavalla harjoitusviikolle kuuden harjoitusviikon jälkeen. Toisaalta tutkimuksessa suoritettiin aerobista harjoittelua, eikä voimaharjoittelua.
Hammarströmin ja kumppanien (2019) erittäin mielenkiintoisessa tutkimuksessa huomattiin korkeamman volyymin kasvattavan enemmän lihasta ja voimaa verrattuna pienempään harjoituskuormaan. Harjoittelu toteutettiin niin, että vastakkainen jalka teki vähemmän volyymia ja toinen jalka enemmän. Harjoituskuorma oli hyvin pieni, mutta ero kolminkertainen (1 sarja vs 3 sarjaa). 13 osallistujaa hyötyi selvästi enemmän suuremmasta volyymista lihasmassan kohdalla ja 16 osallistujaa voiman kohdalla. Ainoastaan kolme osallistujaa hyötyi pienemmästä harjoituskuormituksesta. Lisäksi yksilötasolla korkeampiharjoitusvolyymi oli yhteydessä lisääntyneeseen ribosomien biogeneesiin eli uudimuodostukseen. Myös muut ovat todenneet saman asian (Figueiredo et al. 2015; Stec et al. 2016; Mobley et al. 2018).
Oletko ravi- vai työhevonen?
Lihassolujakauma ei selitä yksin yksilöllisiä eroja voimaharjoittelussa, mutta erittäin mielenkiintoisesti Van Vossel et al. (2023) huomasivat, että saadakseen saman lihasmassan kasvun aikaan enemmän hitaita lihassoluja omaavien pitää tehdä enemmän työtä. Tämä tukee ajatusta siitä, että hidassolukkoisten yksilöiden voi olla kannatavaa tehdä suurempaa volyymiikuormaa. Kannattaa siis rohkeasti kokeilla suurempaa volyymia, jos et kehity. Jotkut yksilöt ovat kuin dieselkoneita, jotka vaativat enemmän kuormaa lähteäkseen kunnolla käyntiin.
Mistä sitten tietää mikä on sopiva harjoituskuorma? Kannattaa aloittaa määrittämällä vähimmäisannos, jolla pystyy kehittymään. Tämän pohjalta voi lähteä etsimään optimaalista harjoituskuormitusannosta (Fisher ym., 2017).
Jos kehitystä ei tule, niin vaihda harjoitusmuotoa
Harjoitusmuodon vaihtamisella voi myös tulla kehitystä. Esimerkiksi tässä mielenkiintoisessa tutkimuksessa 16 rugbypelaajaa tekivät neljä erilaista reeniä, jolla pyrittiin määrittämään testosteronivaste jokaisen eri harjoituksen kohdalla. Tarkoituksena oli mitata, mikä harjoitusmuoto tuotti suurimman ja pienimmän testosteronivasteen. 8 pelaajaa suoritti kolmen viikon harjoitusblokin, jossa tehtiin sitä harjoitusmuotoa, joka tuotti maksimaalisen testosteronivasteen ja toinen porukka suoritti saman pituisen harjoituspätkän, mutta tehden sitä harjoitusmuotoa, joka tuotti pienimmän testosteronivasteen.
Harjoitusliikkeet olivat samat, eli penkki, jalkaprässi, alatalja ja kyyky, mutta sarjat ja toistot vaihtelivat:
Harjoitusmuoto 1: 4 x 10 x 70% 2min palautus.
Harjoitusmuoto 2: 3 × 5 x 85% 3min palautus.
Harjoitusmuoto 3: 5 × 15 x 55% 1min palautus.
Harjoitusmuoto 4: 3 × 5 x 40% 3min palautus.
Kaikki pelaajat kehittyivät merkittävästi penkissä ja jalkaprässissä kun he tekivät harjoitusmuotoa, joka aiheutti suurimman testosteronivasteen. Toisaalta pienimmän hormonaalisen vasteen aiheuttanut harjoitusmuoto ei aiheuttanut mitään muutoksia jopa 75% urheilijoista ja joillakin yhden toiston maksimi jopa pieneni.
Mielenkiintoisesti molemmilla henkilöillä, joilla oli suurin testosteronivaste 3 × 5 x 40% protokollalle, esiintyi 4 × 10 x 70% protokolla vähäisimpänä vasteenaan sekä ennen että jälkeen harjoitusjakson. Samalla tavalla seitsemästä yksilöstä, jotka vastasivat optimaalisesti 4 × 10 x 70% protokollalle, viidelle (71%) 5 × 15 x 55% protokolla tuotti vähiten testosteronivastetta. Tutkimus osoittaa hyvin, että joillekin erilaiset harjoitusmuodot näyttäisivät toimivat paremmin kuin toisille. Onkin tärkeä etsiä juuri yksilölle sopiva harjoitusmuoto.
Henkinen jaksaminen ja vahvuus voivat edesauttaa harjoittelua
Talsnesin ja kumppanien tutkimuksessa pystyttiin erottelemaan korkeasti ja heikosti harjoitteluun vastaavia kestävyysurheilijoita kuuden kuukauden harjoittelujakson jälkeen. Mielenkiintoisesti tutkimuksessa oli haastateltu valmentajia, että mistä erot voisivat johtua. Valmentajien mukaan korkeampi motivaatio ja vahvemmat valmentaja-urheilija suhteet korkean vasteen ryhmässä vaikuttivat yksilöllisesti optimoitujen harjoitus- ja palautumisrutiinien käyttöön, mikä johti positiivisempaan suorituskyvyn kehitykseen. Tämä johti myös siihen, että korkean vasteen ryhmän urheilijat suorittivat suurempia harjoitusmääriä (viikoittainen kuormitus: 3825 ± 1013 vs. 3228 ± 748 ja kuormitus/volyymi-suhde: 4,9 ± 0,6 vs. 4,2 ± 0,5; molemmat P ≤ 0,05) ja heillä oli vähemmän loukkaantumisia/sairastumisia verrattuna heikosti kehittyviin (5 ± 3 vs. 10 ± 5 päivää; P = 0,07).
Yhteenveto
Yksilöiden välillä on runsaasti eroja kehittymisessä
Ei kannata selittää kehittymättömyyttään geeneillä. Geenit asettavat tietyt reunaehdot, mutta kehitys on mahdollista kaikille.
Joillekin yksilöille sopii suurempi harjoituskuorma kuin toisille. Yksilöllisen vasteprofiilin etsiminen on tärkeää.
Erilaisia harjoitusmuotoja kokeilemalla voi myös saada kehitystä aikaan.
Lähteet
Montero,D., and Lundby, C. (2017). Refuting the myth of non-response to exercise training: ’non-responders’ do respond to higher dose of training. J. Physiol. 595, 3377–3387. doi: 10.1113/JP273480
Nunes, J. P., Pina, F. L., Ribeiro, A. S., Cunha, P. M., Kassiano, W., Costa, B. D., et al. (2021b). Responsiveness to muscle mass gain following 12 and 24 weeks of resistance training in older women. Aging Clin. Exp. Res. 33, 1071–1078. doi: 10.1007/s40520-020-01587-z
Scarpelli, M. C., Nóbrega, S. R., Santanielo, N., Alvarez, I. F., Otoboni, G. B., Ugrinowitsch, C., et al. (2020). Muscle hypertrophy response is affected by previous resistance training volume in trained individuals. J. Strength Cond. Res. 27, 1–5. doi: 10.1519/JSC.0000000000003558
Fisher, J. P., Steele, J., Gentil, P., Giessing, J., andWestcott,W. L. (2017). A minimal dose approach to resistance training for the older adult; the prophylactic for aging. Exp. Gerontol., 99, 80–86. doi: 10.1016/j.exger.2017.09.01
Kramer, J. B. (1997). Effects of single vs multiple sets of weight training: impact of volume, intensity, and variation. J. Strength Cond. Res. 11, 143–147. doi: 10.1519/00124278-199708000-00002
Radaelli, R., Botton, C. E., Wilhelm, E. N., Bottaro, M., Brown, L. E., Lacerda, F., et al. (2014a). Time course of low- and high-volume strength training on neuromuscular adaptations and muscle quality in older women. Age 36, 881–892. doi: 10.1007/s11357-013-9611-2
Radaelli, R.,Wilhelm, E. N., Botton, C. E., Rech, A., Bottaro,M., Brown, L. E., et al. (2014b). Effects of single vs. multiple-set short-term strength training in elderly women. Age 36:9720. doi: 10.1007/s11357-014-9720-6
Krieger, J. W. (2010). Single vs. multiple sets of resistance exercise for muscle hypertrophy: a meta-analysis. J. Strength. Cond. Res. 24, 1150–1159. doi: 10.1519/JSC.0b013e3181d4d436
Jones, N., Kiely, J., Suraci, B., Collins, D. J., de Lorenzo, D., Pickering, C., … & Grimaldi, K. A. (2016). A genetic-based algorithm for personalized resistance training. Biology of sport, 33(2), 117.
Eynon, N., Hanson, E. D., Lucia, A., & Houweling, P. J. (2013). Genes for elite power and sprint performance: ACTN3 leads the way. Sports Medicine, 43(9), 803-817.
Rhea, M. R., Alvar, B. A., Burkett, L. N., & Ball, S. D. (2003). A meta-analysis to determine the dose response for strength development. Medicine and science in sports and exercise, 35(3), 456-464.
Phillips, S. M., & Van Loon, L. J. (2011). Dietary protein for athletes: from requirements to optimum adaptation. Journal of sports sciences, 29(sup1), S29-S38.
Burd, N. A., Holwerda, A.M., Selby, K. C.,West, D.W., Staples, A.W., Cain, N. E., et al. (2010a). Resistance exercise volume affects myofibrillar protein synthesis and anabolic signalling molecule phosphorylation in young men. J. Physiol. 588, 3119–3130. doi: 10.1113/jphysiol.2010.192856
Terzis, G., Spengos, K., Mascher, H., Georgiadis, G., Manta, P., and Blomstrand, E. (2010). The degree of p70 S6k and S6 phosphorylation in human skeletal muscle in response to resistance exercise depends on the training volume. Eur. J. Appl. Physiol. 110, 835–843. doi: 10.1007/s00421-010-1527-2
Garma, T., Kobayashi, C., Haddad, F., Adams, G. R., Bodell, P. W., and Baldwin, K. M. (2007). Similar acute molecular responses to equivalent volumes of isometric, lengthening, or shortening mode resistance exercise. J. Appl. Physiol. 102, 135–143. doi: 10.1152/japplphysiol.00776.2006
Peterson,M. D., Sen, A., and Gordon, P.M. (2011). Influence of resistance exercise on lean body mass in aging adults: a meta-analysis. Med. Sci. Sports Exerc. 43, 249–258. doi: 10.1249/MSS.0b013e3181eb626
Hammarström, D., Øfsteng, S., Koll, L., Hanestadhaugen, M., Hollan, I., Apró, W., Whist, J.E., Blomstrand, E., Rønnestad, B.R. and Ellefsen, S. (2020), Benefits of higher resistance-training volume are related to ribosome biogenesis. J Physiol, 598: 543-565. https://doi.org/10.1113/JP278455
Figueiredo VC, Caldow MK, Massie V, Markworth JF, Cameron-Smith D & Blazevich AJ (2015). Ribosome biogenesis adaptation in resistance training-induced human skeletal muscle hypertrophy. Am J Physiol Endocrinol Metab 309, E72– E83.
Mobley CB, Haun CT, Roberson PA, Mumford PW, Kephart WC, Romero MA, Osburn SC, Vann CG, Young KC, Beck DT, Martin JS, Lockwood CM & Roberts MD (2018). Biomarkers associated with low, moderate, and high vastus lateralis muscle hypertrophy following 12 weeks of resistance training. PLoS One 13, e0195203.
Stec MJ, Kelly NA, Many GM, Windham ST, Tuggle SC & Bamman MM (2016). Ribosome biogenesis may augment resistance training-induced myofiber hypertrophy and is required for myotube growth in vitro. Am J Physiol Endocrinol Metab 310, E652– E661.
Beaven, C Martyn1; Cook, Christian J2; Gill, Nicholas D1. Significant Strength Gains Observed in Rugby Players after Specific Resistance Exercise Protocols Based on Individual Salivary Testosterone Responses. Journal of Strength and Conditioning Research 22(2):p 419-425, March 2008. | DOI: 10.1519/JSC.0b013e31816357d4
Talsnes RK, van den Tillaar R, Cai X, Sandbakk Ø. Comparison of High- vs. Low-Responders Following a 6-Month XC Ski-Specific Training Period: A Multidisciplinary Approach. Front Sports Act Living. 2020 Sep 8;2:114. doi: 10.3389/fspor.2020.00114. PMID: 33345103; PMCID: PMC7739740.
Bathgate KE, Bagley JR, Jo E, Talmadge RJ, Tobias IS, Brown LE, Coburn JW, Arevalo JA, Segal NL, Galpin AJ. Muscle health and performance in monozygotic twins with 30 years of discordant exercise habits. Eur J Appl Physiol. 2018 Oct;118(10):2097-2110. doi: 10.1007/s00421-018-3943-7. Epub 2018 Jul 14. PMID: 30006671.
Marsh CE, Thomas HJ, Naylor LH, Scurrah KJ, Green DJ. Fitness and strength responses to distinct exercise modes in twins: Studies of Twin Responses to Understand Exercise as a THerapy (STRUETH) study. J Physiol. 2020 Sep;598(18):3845-3858. doi: 10.1113/JP280048. Epub 2020 Jul 7. PMID: 32567679.
Davidsen PK, Gallagher IJ, Hartman JW, Tarnopolsky MA, Dela F, Helge JW, Timmons JA, Phillips SM. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J Appl Physiol (1985). 2011 Feb;110(2):309-17. doi: 10.1152/japplphysiol.00901.2010. Epub 2010 Oct 28. PMID: 21030674.
Van Vossel, K., Hardeel, J., de Casteele, F.V., Van der Stede, T., Weyns, A., Boone, J., Blemker, S., Lievens, E. and Derave, W. (2023), Can muscle typology explain the inter-individual variability in resistance training adaptations?. J Physiol. Accepted Author Manuscript. https://doi.org/10.1113/JP284442
Figueiredo VC, de Salles BF, Trajano GS. Volume for Muscle Hypertrophy and Health Outcomes: The Most Effective Variable in Resistance Training. Sports Med. 2018 Mar;48(3):499-505. doi: 10.1007/s40279-017-0793-0. PMID: 29022275.
Mattocks KT, Buckner SL, Jessee MB, Dankel SJ, Mouser JG, Loenneke JP. Practicing the Test Produces Strength Equivalent to Higher Volume Training. Med Sci Sports Exerc. 2017 Sep;49(9):1945-1954. doi: 10.1249/MSS.0000000000001300. PMID: 28463902.
Ensimmäisessä osassa pureuduttiin isometrisen harjoittelun teoriaam ja isometrisen kuormituksen aiheuttamiin adaptaatioihin. Tässä toisessa osassa syvennytään ohjelmointiesimerkkeihin ja käydään myös erilaisia liike-esimerkkejä läpi isometrisestä voimaharjoittelusta. Isometrisessä harjoittelussa vain mielikuvitus toimii rajoittavana tekijänä harjoitevalinoissa.
Käytä räjähtävää tai ballistista voimantuottotapaa isometrisessä harjoittelussa
Isometrinen voimaharjoittelu voi parantaa voimantuottonopeutta, mutta vain jos se tehdään räjähtävästi. Nopea voimantuotto on parantunut erityisesti räjähtävällä tai ballistisella voimaharjoittelulla (1, 2 & 3). Kun tavoitteena on nopean voimantuoton parantaminen, näyttäisi intentio olevan yhtä tärkeä kuin ulkoinen supistus, koska haluttu muutos on joka tapauksessa hermostollinen ei lihastyötavalla ole väliä (4). Joten käytä isometrisessä voimaharjoittelussa nopeaa voimantuottoa.
Isometrinen polven yläpuolelta tehtävä veto on klassinen isometrinen voimaharjoittelu- ja mittausmetodi.
Bogdanis ja kumppanit (6) huomasivat, että isometrinen harjoittelu paransi dynaamista maksimikyykkyä kymmenisen prosenttia ja esikevennettyä hyppyä noin seitsemän prosenttia isometrisen jalkaprässiharjoittelun jälkeen. Isometrinen voimaharjoittelun siirtovaikutus on hyvä yksinkertaisiin liikkeisiin, mutta kompleksisemmat liikkeet vaativat enemmän koordinaatioharjoittelua kehittyäkseen. Isometrinen harjoittelu ei välttämättä haasta motorista aivokuorta tarpeeksi ja motorinen oppiminen saattaa jäädä pienemmälle roolille verrattuna dynaamiseen harjoitteluun. Tämä kannattaa huomioida harjoittelua suunnitellessa.
Askelkyykky pinnoilta on hyvä tapa harjoitella kulmaspesifisti nopeaa voimantuottoa.
Isometrisen harjoittelun hyödyt nopeuslajeissa ja pikajuoksijalle?
Urheilussa tärkeä ominaisuus on se, että miten nopeasti voimaa pystytään tuottamaan. Urheilu tapahtuu usein sekunnin kymmenyksissä ja sadasosissa ja näissä lyhyissä hetkissä pitäisi pystyä tuottamaan mahdollisimman paljon voimaa liikkuakseen eteenpäin, vaihtaakseen suuntaan, ponnistaakseen tai esimerkiksi heittääkseen välinettä. Voimantuottonopeuden kehittäminen ilman suurempaa metabolista väsymystä on isometrisen harjoittelun yksi etu verrattuna dynaamiseen harjoitteluun.
Konsentrinen työvaihe on perinteisen voimaharjoittelun rajoittava lihastyötapa. Perinteisen kyykyn tai maastavedon aikana isometrinen ja eksentrinen lihastyötapa ei tule tarpeeksi kuormitetuksi. Vaikka liikkeessä onkin eksentrinen ja isometrinen osuus, niin ne ovat submaksimaalisia vaiheita. Jos haluaa kuormittaa perinteisen voimaharjoittelun keinoin isometristä tai eksentristä lihastyötapaa, niin pitää liikettä hidastaa tai lisätä stoppeja liikkeeseen. Tämä on hyvä keino, mutta lisää huomattavasti liikkeen metabolista rasitusta, eikä niinkään liikkeen hermostollista osuutta. Lisääntynyt väsymys ja vähentynyt tehontuotto ei ole tavoiteltavia asioita nopeuslajin urheilijoille, ainakaan kilpailukaudella. Lisäksi perinteinen voimaharjoittelu aiheuttaa mekaanista vaurioita, josta toipumiseen menee aikaa. Isometrinen harjoittelu näyttäisi pitävän urheilijan hiukan tuoreempana, jotta nopeusharjoittelua voidaan tehdä mikrosyklin sisällä yhdessä voimaharjoittelun kanssa.
Takareisipidot ovat loistava tapa kuormittaa pitkällä lihaspituudella takareiden isometristä voimantuottoa.
Isometrisen harjoittelun hyödyt piilevät nopeuslajien urheilijoille siinä, että ne voidaan suorittaa maksimaalisella intensiteetillä ilman kovaa väsymystä. Tämä johtuu suurelta osin eksentrisen vaiheen puutteesta, joten lihasvaurioita ei pääse samassa määrin syntymään. Lisäksi pienempi metabolinen rasitus isometrisessä harjoittelussa vähentää urheilijan akuuttia väsymystä.
Miten toteuttaa isometristä voimaharjoittelua käytännössä
Isometrisen harjoittelussa törmätään yleensä ongelmaan, että miten progressoida harjoittelua. Voiman kasvua voi mitata voimalevyillä tai erilaisilla venymämittareilla. Ilman näitä on vaikea havainnoida tuottaako urheilija tosissaan maksimaalisen määrän voimaa toiston aikana. Koska nopeuslajin urheilijoille tavoitteena on aina tuottaa maksimimäärä voimaa ja vielä mahdollisimman nopeasti, on intensiteettiprogressio haastavaa. Volyymiprogressio on taas tehokas tapa edetä, mutta sen kanssa pitää olla erittäin maltillinen. Harjoitusfrekvenssin lisääminen on toinen tehokas tapa lisätä volyymia. Viikossa tulisi olla noin 40- 60 s nopeaa voimantuottoon tähtäävää isometristä harjoittelua, jos tavoitteena on nopean voimantuoton parantaminen. Progressio voi olla esimerkiksi taulukko 1:den kaltainen.
Harjoituskerrat
Yhden harjoituskerran volyymi (s)
Kokonaisvolyymi (s)
Viikko 1
2
20
40
Viikko 2
2
25
50
Viikko 3
2
30
60
Viikko 4
3
20
60
Viikko 5
3
25
75
Viikko 6
3
30
90
Taulukko 1: Yli kuuden viikon ohjelmia ei kannata nopeassa isometrisessä voimantuottoharjoittelussa tehdä. Kuden viikon jälkeen tarvitaan viimeistään ärsykkeen vaihtelua.
Kuten kaikki nopeusharjoittelu pitäisi myös räjähtävää isometristä voimaharjoittelua tehdä tuoreena ja levänneen, jotta voidaan maksimoida nopea voimantuotto. Nopea voimantuotto näyttäisi laskevan jopa viiden toiston jälkeen (6), joten suositeltavaa olisi pitää toistomäärä vähäisinä (1-5). Sarjaprogressio määrässä on tehokkaampi tapa kuin toistojen lisääminen, eli esimerkiksi neljästä sarjasta kohti kymmentä sarjaa. Tärkeintä on kuitenkin, että harjoittelu suoritetaan maksimaalisella intentiolla.
Esimerkiksi Olsen ja Hopkins (7) laittoivat huippukamppailulajiurheilijat tekemään lajiliikespesifiä isometristä harjoittelua. Kamppailijat tekivät räjähtävää isometristä potkuliikettä, kun potkua suorittava jalka oli sidottu vyöllä kiinni ylös. Kamppailijat tekivät neljä sarjaa kymmenen toistoa (muutama sekunti) yhdeksän viikon ajan. Ohejlmassa oli maltillinen volyymiprogressio. Tutkijat huomasivat jopa 11-21 prosentin kasvun liikenopeudessa eri potkuliikkeissä.
Miten progressoida harjoittelua kuuden viikon ohjelman jälkeen?
Haluaisitko integroida isometrisen voimaharjoittelun mukaan ohjelmaan koko vuodeksi? Yksi hyvä tapa on tehdä intensiteettiprogressio ohjelmasta toiseen lisäämällä vauhtia. Esimerkiksi tämän kaltaisella ohjelmalla:
Intensiteetin kehittäminen. Vauhdin lisääminen blokista toiseen.
Isometrinen maksimaalinen työ liikkumatonta objektia vastaan tavoitteena kehittää spesifiä hypertrofiaa ja vahvistaa jänteitä (8-12 viikoa).
Räjähtävät isometriset (esim taulukon 1 ohjelma, noin 6 vikkoa)
Isometriset vaihdot (6 viikkoa)
Isometriset kiinniotot (6 viikkoa).
Isometriset vaihdot tarkoitavat dynaamisen liikkeen lisäämistä isometriseen voimaharjoitteluun. Tässä on tarkoitus haastaa isometristä voimaharjoittelua lisäämällä raajaan liikenopeutta, mikä pitää isometrisen voimaharjoittelun aikana pysäyttää ja hallita. Tämän kaltaista lihastyötä tehdään urheilussa suorituksissa huomattavan paljon. Alla muutama esimerkki isometrisistä vaihdoista, mutta jälleen vain mielikuvitus on isometrisessä voimaharjoittelussa rajana. Mikä liike palvelisi lajisi suorituskykyä parhaimmalla mahdollisella tavalla?
Isometriset kiinniotot taas ovat isometrisistä vaihdoista seuraava vauhdikkaampi askel. Näissä lisätään yhä enemmän raajan vauhtia. Mukaan tulee myös eksentrinen osuus ennen isometristä osuutta, joten kokonaisuudessaan nämä ovat jo hyvin lähellä dynaamisia liikkeitä.
Yhteenveto
Isometrinen harjoittelu on loistava tapa kehittää nopeaa voimantuottoa ilman suurempaa metabolista rasitusta. Toimii erityisen hyvin kilpailukauden aikana. Mekanismit nopean isometrisen voimantuoton ja hitaan isometrisen voimantuoton välillä vaihtelevat huomattavasti.
Esimerkiksi Maffiuletti ja Marin (8) vertailivat isometristä jalkaprässiä niin, että toinen ryhmä teki sitä räjähtävästi 1s ajan (yritti tuottaa mahdollisimman paljon voimaa mahdollisimman nopeasti) ja toinen ryhmä progressiivisesti 4s ajan (voimaa lisättiin rauhallisesti suorituksen toiston ajan lisää). Kummatkin ryhmät paransivat voimantuottoa huomattavasti. Mekanismit taustalla vaihtelivat, kun progressiivinen hitaampi isometrinen harjoittelu vaikutti vastus lateraliksen M-aaltojen ominaisuuksiin ilman vaikuttamatta lihassoluihin. M-aalto kuvastaa kaikkien motoristen yksiköiden yhtäaikaista syttymistä ja on kaikkien aktiopotentiaalien summa. Lyhyempi räjähtävä voimantuotto vaikutti lihassolujen supistuvien osien ominaisuuksiin, kun taas M – aallossa ei havaittu mitään muutoksia.
Onkin tärkeä tietää mitä haluaa kehittää ja miksi. Pelkkä isometrisen harjoittelun trendikkyys ei riitä syyksi ruveta tekemään sitä.
Alla listattuna muutamia erilaisia isometrisia harjoitteita;
Balshaw TG, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specific functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol. 2016;120(11):1364-1373.
Tillin NA, Folland JP. Maximal and explosive strength training elicit distinct neuromuscular adaptations, specific to the training stimulus. Eur J Appl Physiol. 2014;114(2):365-374.
Maffiuletti NA, Martin A. Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Med Sci Sports Exerc. 2001;33(7):1220-1227
Behm DG, Sale DG. Intended rather than actual movement velocity determines velocity-specific training response. J Appl Physiol. 1993;74(1):359-368.
Bogdanis GC, Tsoukos A, Methenitis SK, Selima E, Veligekas P, Terzis G. Effects of low volume isometric leg press complex training at two knee angles on force-angle relationship and rate of force development. Eur J Sport Sci. 2018;1-9. https://doi.org /10.1080/17461391.2018.1510989. [Epub ahead of print].
Viitasalo JT, Komi PV (1981) Effects of fatigue on isometric force- and relaxation-time characteristics in human muscle. Acta Physiologica Scandavica 111(1):87–95.
Olsen PD, Hopkins WG (2003) The effect of attempted ballistic training on the force and speed of movements. Journal of Strength and Conditioning Research 17(2):291–98.
Maffiuletti NA, Martin A (2001) Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Medicine and Science in Sports and Exercise 33(7):1220–27.
Isometrisellä lihastyötavalla viitataan lihastyöhön, missä lihasjännekompleksin pituudessa ei tapahdu muutosta. Konsentrisessa lihastyötavassa lihas lyhenee supistuessaan ja eksentrisessä lihastyötavassa lihas pitenee lihassolujen supistuessa. Esimerkkinä kyykystä ylös ponnistaminen on konsentrista lihastyötä suurimmalla osalla jalkojen lihaksia ja lihasten pituus lyhenee, kun taas eksentrinen toiminta pidentää lihaspituutta. Tästä esimerkkinä, kun mennään alaspäin kyykyssä. Isometristä harjoittelua voidaan käytännössä tehdä monella eri tavalla, mutta tässä jutussa keskitytään pelkästään työskentelyyn liikkumatonta esinettä vasten.
Isometrinen harjoittelu on erityisen mielenkiintoinen aihealue urheilijoille, koska isometristä harjoittelua voi hyödyntää positiivisten hermolihasjärjestelmän adaptaatioiden saavuttamiseksi ilman liiallista väsymystä, mitä perinteinen keskiraskas voimaharjoittelu aiheuttaa.
VIDEO 1: Yleisin tapa toteuttaa käytännössä isometristä voimaharjoittelua liikkumatonta esinettä vasten on käyttää tankoa ja suojarautoja tai vastaavia tukirakenteita.
Minkälaisia adaptaatiota isometrinen harjoittelu aiheuttaa
Voiko pelkällä isometrisellä harjoittelulla kasvattaa lihasta?
Kyllä voi! Isometrinen harjoittelu 42–100 päivän ajan on johtanut 5,4–23% lihaksen poikkipinta-alan kasvuun ja jopa 91,7% nousuun maksimivoimassa (28-37). Pidempikestoinen interventio näyttäisi vaikuttavan huomattavasti lihaksen kokoon. Mitä pidempi interventio oli, sitä enemmän lihas kasvoi. Hypertrofiaan vaikutti myös harjoittelun intensiteetti, voluumi, supistuksen kesto ja lihaksen pituus.
Erityisesti pitkillä lihaspituuksilla tehty isometrinen harjoittelu parantaa ylivoimaisesti enemmän lihaksen kokoa verrattuna lyhyillä lihaspituuksilla tehtyyn isometriseen harjoitteluun, vaikka volyymi olisi tasattu näiden ryhmien välillä (1, 2 & 3). Tulokset ovat lähes samansuuntaisia, kun verrataan isometristä harjoittelua dynaamiseen harjoitteluun. Myös normaalissa dynaamisessa voimaharjoittelussa näyttäisi laaja liikerata olevan huomattavasti hyödyllisempi hypertrofian kannalta verrattuna vajaisiin liikeratoihin (4, 5 & 6). Yksi syy tähän voi olla, että pitkällä lihaspituudella tehdyt supistukset näyttäisivät tuottavan huomattavasti enemmän lihasvaurioita verrattuna lyhyellä lihaspituudella tehtäviin harjoitteisiin (7). Tämä johtuu siitä, että nivelen vipuvarsi kasvaa pitkillä lihaspituuksilla ja näin lisää mekaanista jännitystä lihaksessa verrattuna lyhyempään vipuvarteen. Suurempi mekaaninen jännitys aiheuttaa enemmän lihasvaurioita. Lisäksi pitkät lihaspituudet kuluttavat enemmän happea, vaativat enemmän verenkierrolta töitä ja kokonaisuudessaan lisäävät metaboliittien kerääntymistä enemmän kuin lyhyet lihaspituudet (8). Metaboliset tekijät ovat tutkitusti myös yhteydessä lihaskasvuun (9). Eli jos tavoitteena on spesifi lihaskasvu isometrisessä harjoittelussa, niin pitkät lihaspituudet ovat ehdottomasti paras valinta.
Volyymillä on selvästi väliä myös isometrisessä harjoittelussa, kun tavoitteena on lihaskasvu. Meyers (10) vertaili matala volyymista harjoittelua (3 x 6 sekuntia maksimaalisella intensiteetillä) korkea volyymiseen harjoitteluun (20 x 6 sekuntia maksimaalisella intensiteetillä) hauislihaksella. Kuuden viikon jälkeen enemmän volyymia tehnyt ryhmä oli saavuttanut selvästi isomman muutoksen hauislihaksen ympärysmitassa verrattuna matalavolyymiseen ryhmään. Myös Balshaw ja kumppanit (11) totesivat, että suurempi määrä volyymia (40 x 3 sekuntia 75% isometrisestä maksimista) tuotti enemmän lihaskasvua etureiteen 12-viikon aikana verrattuna pienempään harjoituskuormaan (40 x 1 sekuntia 80% isometrisestä maksimista).
Mielenkiintoista on myös, että Schott ja kumppanit (12) löysivät, että pidempikestoinen harjoittelu (4 x 30 sekuntia) tuotti enemmän hypertrofiaa verrattuna lyhyempikestoiseen harjoitteluun (4 x 10 x 3 sekuntia), vaikka liikesuoritteiden kokonaiskesto oli lopulta sekunnilleen yhtä pitkä. 14-viikon harjoittelun jälkeen etureiden vastus lateralis lihas kasvoi jopa 11,1% enemmän, kun lyhyempikestoisia supistuksia tehneellä ryhmällä ei löydetty ollenkaan merkitsevää muutosta etureiden kasvusta! Tämä voi johtua siitä, että pitkään ylläpidetyt supistukset estävät verenkierron ja vähentävät hapen saturaatiota alueella, stimuloiden näin hypertrofiaa monien paikallisten ja systeemisten mekanismien kautta.
KUVA 1: Isometristä harjoittelua voidaan tutkia nilkan plantaari- tai dorsifleksiota tarkkailemalla tämän näköisellä koeasetelmalla.
Isometrinen harjoittelu muokkaa myös lihaksen arkkitehtuuria
Hypertrofiaa haettaessa lihastyötavalla ei ole hirveästi merkitystä, sillä niin dynaamisella, eksentrisellä ja isometrisellä harjoittelulla voidaan saada lihaskasvua aikaiseksi, mutta jos tavoitteena on saada muutoksia aikaan lihaksen arkkitehtuuriin, on lihatyötavalla todellakin merkitystä.
Laadukkaita tutkimuksia aiheesta ei ole paljoa, joten päätelmien tekeminen on haastavaa, mutta Noorkoiv ja kumppanit (3) huomasivat, että pidemmällä lihaspituudella tehty isometrinen harjoittelu (polvikulma 38.1 ± 3.7°) kasvatti vastus lateraliksen lihasfasciculuksen (lihassolukimppu, jota ympäröi lihaskalvo) pituutta keskiosassa lihasta merkitsevästi. Mielenkiintoisesti lyhyemmällä lihaspituudella tehty harjoittelu kasvatti taas distaalisessa päässä olevan lihasfasciculuksen pituutta. Ainoastaan yksi toinen tutkimus (1) on raportoinut vastus lateraliksen lihasfasciculuksen pituuden lisääntymistä ja myös pennaatiokulman muutoksesta pitkällä lihaspituudella tehdyn isometrisen harjoittelun jälkeen.
Isometrinen voimaharjoittelu näyttäisi aiheuttavan muutoksia lihaksen arkkitehtuuriin ja erityisesti lisäävän lihasfasciculuksen pituutta ja kenties jopa aiheuttaa muutoksia pennaatiokulmaan. Tällä on erityisesti väliä, jos tavoitteena on tehdä urheilijoista nopeampia, sillä esimerkiksi sprinttereillä on pidemmät lihasfasciculukset jaloissa verrattuna kestävyysurheilijoihin (38) ja 100 metrin juoksusuoritus on yhdistetty lihasfasciculuksien pituuksiin (39).
Isometrisen harjoittelun vaikutukset jänteisiin
Jänteen tarkoitus on siirtää voimia luun ja lihaksen välillä mahdollistaen nivelen liike. Ennen ajateltiin jänteiden olevan muuttumattomia, mutta onneksi nykyään tiedetään jo, että jänteet kykenevät adaptoitumaan stimulukseen merkitsevästi ja voivat käydä todella isoja arkkitehtuurisia muutoksia läpi pitkäaikaisen kuormituksen johdosta.
Esimerkiksi kun vertaillaan eri lajien urheilijoita akillesjännerepeämän kokemiin ihmisiin, on huomattu, että esimerkiksi lentopalloilijoilla on huomattavasti suurempi akillesjänne (119 ± 5.9) verrattuna akillesjännerepeämän kokemiin ihmisiin (101 ± 5.4). Mielenkiintoista oli, että kajakkiurheilijoilla oli lähes samankokoinen akillesjänne kuin repeämän kokemilla ihmisillä (101 ± 5.6) (13). Kajakkiurheilijat eivät juuri käytä akillesjänteitään lajissaan, joten harjoittelulla näyttäisi olevan suuri vaikutus jänteen rakenteisiin.
Jänteen adaptaatiot ovat erittäin tärkeitä ja haluttuja adaptaatioita nopeuslajin urheilijoille, sillä jänne toimii nopeassa liikkeessä liikuttajana jousen tavoin. Intensiteetti on ehdottomasti tärkein muuttuja jänteen adaptaatioissa. Kova intensiteettinen isometrinen plantaarifleksion harjoittelu (noin 90 % isometrisestä maksimista) lisäsi akillesjänteen poikkipinta-alaa ja jäykkyyttä 14-viikon harjoitteluohjelman aikana jopa parhaimmillaan 36 % (14 & 15). Samaa ei huomattu matalaintensiteettisellä harjoittelulla (55 % isometrisestä maksimista). Myös muut ovat raportoineet samankaltaisia runsaita muutoksia jänteen jäykkyydessä (vaihteluväli 17,5 % - 61,6 %) isometrisen voimaharjoittelun seurauksena intensiteetin vaihdellessa 70–100 % välillä isometrisestä maksimivoimasta (16, 17 & 18). Näyttäisi siltä, että 70 % voisi olla minimi-intensiteetti, joka vaaditaan haluttujen jänneadaptaatioiden saavuttamiseksi.
Räjähtävä isometrinen voimaharjoittelu taas lisäsi jänteen aponeuroosin elastisuutta, mutta vähensi jänteen poikkipinta-alaa (-2,8 %) (19). Isometrisen harjoittelun intensiteetillä ja kestolla saavutetaan hyvin erilaisia adaptaatioita. Jänteiden vahvistamisessa tulee suosia pidempiä ja intensiteetti korkealla tehtyjä supistuksia, kun taas kisakaudella voi tehdä terävämpiä elastisuutta lisääviä erittäin lyhyitä supistuksia. Lisäksi pidempi lihaspituus näyttäisi kehittävän jänteen jäykkyyttä enemmän kuin harjoittelu lyhyellä lihaspituudella samalla tavalla kuin lihaskasvussa (2).
KUVA 2: Bruce Leekin käytti isometristä harjoittelua kehittämään omaan suorituskykyään.
Isometrisen voimaharjoittelun vaikutukset hermostoon
Hermoston adaptaatiot ovat kokonaisuudessaan hyvin harjoitteluspesifejä. Esimerkiksi Balshaw ja kumppanit (11) vertailivat 12 viikon aikana maksimaalista voimaharjoittelua (1 sekunnin rauhallinen nousu 75% isometrisestä maksimista ja siellä 3s pito) räjähtävään voimaharjoitteluun (mahdollisimman nopeasti >90% isometriseen maksimiin ja siellä 1s pito). Isometrinen maksimivoima kehittyi eniten maksimivoimaharjoittelulla, mutta räjähtävä voimaharjoittelu lisäsi EMG aktiivisuutta ihan liikkeen alussa (0–100 ms aikana) enemmän verrattuna maksimivoimaharjoitteluun. Nämä adaptaatiot olivat hermostoperäisiä ja olivat harjoitteluspesifejä, kun maksimivoimaharjoittelu kehitti maksimivoimaa ja räjähtävä voima kehitti nopeaa voimantuottokykyä. Myös ballistinen isometrinen harjoittelu on johtanut samankaltaisiin tuloksiin ja EMG amplitudin paranemiseen ensimmäisen 0-150 ms aikana verrattuna maksimivoimaharjoitteluun (11, 23 & 24).
Isometrisellä voimaharjoittelulla voidaan vaikuttaa lihaksen jännitys-pituussuhteeseen, eli siihen, millä lihaksenpituudella tai nivelen kulmalla tuotetaan isoin mahdollinen voima. Tämä on erityisen tärkeä urheilussa, jossa halutaan maksimoida suurin mahdollinen tuotettu voima halutussa asennossa. Myös parasta voimantuottokulmaa voidaan muokata isometrisellä harjoittelulla. Esimerkiksi Alegre ja kumppanit (25) raportoivat, että pidemmällä lihaspituudella harjoittelu kahdeksan viikon ajan johti 11 asteen muutokseen kohti pidempiä lihaspituuksia, kun taas lyhyemmillä kulmilla harjoittelu johti 5,3 astetta optimaalista kulmaa toiseen suuntaan. Myös Bogdanis ja kumppanit (26) huomasivat noin 10 % tiputuksen optimaalisessa kulmassa lyhyillä liikeradoilla harjoitellessa.
Pidempikestoinen supistus näyttäisi olevan joissakin tapauksissa tehokkaampi tapa parantaa voimaa ja myös dynaamista urheilun suorituskykyä (hyppäämistä ja juoksemista) verrattuna nopeaan isometriseen voimantuottotapaan (40). Pidemmässä supistuksessa tehtiin kolmen sekunnin ajan työtä ja räjähtävässä nopeassa isometrisessä voimantuottotavassa tehtiin yhden sekunnin verran töitä. Tuloksia on tulkittava hieman varovasti, sillä pidempää supistusta tehnyt ryhmä teki yhteensä 15 sekunnin verran työtä sarjassa, kun lyhyempää pätkää tehnyt ryhmä teki vain 10 sekunnin verran työtä. Kuuden viikon aikana ja 12 harjoituskerran vuoksi erot kertaantuvat ja tehty kokonaistyö oli huomattavasti isompi kolmen sekunnin ryhmässä verrattuna yhden sekunnin ryhmään. Tämä varmasti osaltaan selittää tuloksia.
Kolmen sekunnin ryhmä paransi esikevennettyä hyppyä 12,1 % ja yhden sekunnin ryhmä 10,8 %. Erot kasvaneista voimatasoissakin voivat selittää nämä muutokset. Mielenkiintoisesti pidempikestoinen isometrinen voimaharjoittelu aiheutti 1,4 % parannuksen 30 metrin juoksuaikaan. Tässäkin tapauksessa enemmän harjoitellut ryhmä paransi huomattavasti enemmän nopeuttaan, kun vähemmän harjoitellut ryhmä. Voisiko kasvaneet voimatasot, ei niinkään nopeus, selittää erot. Normaalilla kovalla kyykyllä ja plyometrisellä harjoittelulla on saatu 1,2 % parannus 30 metrin juoksuaikaan (43), joka on aika lähellä tämän tutkimuksen saamia tuloksia.
Toisaalta tässäkin tutkimuksessa huomattiin, että kyky tuottaa voimaa nopeammin parani yhden sekunnin ryhmällä enemmän kuin kolmen sekunnin ryhmällä, kun taas pidempikestoisessa supistuksessa maksimivoima kehittyi enemmän. Myös muut ovat raportoineet samankaltaisia tuloksia (41 & 42).
Yhteenveto
Isometristä harjoittelua voi hyödyntää positiivisten hermolihasjärjestelmän adaptaatioiden saavuttamiseksi ilman liiallista väsymystä. Tämä on erityisen tärkeää erityisesti urheilijoilla kilpailukauden aikana. Lisäksi jos tiettyä voimantuottokulmaa tai lajin vaatimia kulmia pitää harjoitella, niin isometrinen harjoittelu on erittäin tehokas työkalu niihin.
Isometrinen harjoitteluun pätee samat lainalaisuudet kuin muuhunkin harjoitteluun. Hypertrofiaa saavuttaaksesi tulee harjoittelua tehdä 70-75% intensiteetillä maksimaalisesta supistuksesta noin 3-30s ajan toistossa ja sarjamäärän ollessa > 80 – 150s per yksi harjoituskerta. Maksimivoimaa saavuttaaksesi isometristä harjoittelua tulee tehdä 80-100% maksimaalisesta supistuksesta 1-5s ajan ja kokonaiskeston ollessa 30-90s. Voimantuottonopeutta parantaakseen tulee suorituksessa pyrkiä tuottamaan mahdollisimman nopeasti mahdollisimman paljon voimaa. Sarjan keston tulee olla lyhyt. Kuvassa 3 on koottu tämänhetkiseen tutkimusnäyttöön perustuen ohjeistus isometriseen voimaharjoitteluun.
KUVA 3: Tämänhetkisestä tukimusnäytöstä koostettu taulukko miten isometristä harjoittelua tulisi tehdä, jos haluaa saavuttaa tietyn adaptaation.
Tiivistys
Lihastakin voin kasvattaa pelkällä isometrisellä harjoittelulla. Volyymi ja lihaspituus ovat tärkeimmät muuttujat, kun tavoitteena on lihaskasvu.
Isometrisessä harjoittelussa intensiteetti on päämuuttuja voiman kohdalla. Hypertrofian kohdalla volyymi.
Ballistisella protokola on ylivoimainen räjähtävän voiman kehittymiseen. Ensimmäiselle 50 ja 100 ms voi parantaa voimantuottoa huomattavasti. Jos tämä on tavoite, niin harjoitteet tulisi tehdä mahdollisimman nopeasti ja mahdollisimman voimakkaasti.
Lähteet:
Alegre LM, Ferri-Morales A, Rodriguez-Casares R, Aguado X. Effects of isometric training on the knee extensor moment–angle relationship and vastus lateralis muscle architecture. Eur J Appl Physiol. 2014;114(11):2437-2446.
Kubo K, Ohgo K, Takeishi R, et al. Effects of isometric trainingmat different knee angles on the muscle–tendon complex in vivo. Scand J Med Sci Sports. 2006;16(3):159-167.
Noorkoiv M, Nosaka K, Blazevich AJ. Neuromuscular adaptations associated with knee joint angle-specific force change. Med Sci Sports Exerc. 2014;46(8):1525-1537.
Guex K, Degache F, Morisod C, Sailly M, Millet GP. Hamstring architectural and functional adaptations following long vs. short muscle length eccentric training. Front Physiol. 2016;7(340):1-9.
Barak Y, Ayalon M, Dvir Z. Transferability of strength gains from limited to full range of motion. Med Sci Sports Exerc. 2004;36(8):1413-1420.
Massey CD, Vincent J, Maneval M, Moore M, Johnson JT. An analysis of full range of motion vs. partial range of motion training in the development of strength in untrained men. J Strength Cond Res. 2004;18(3):518-521.
Allen TJ, Jones T, Tsay A, Morgan DL, Proske U. Muscle damage produced by isometric contractions in human elbow flexors. J Appl Physiol. 2018;124(2):388-399.
de Ruiter CJ, de Boer MD, Spanjaard M, de Haan A. Knee angle-dependent oxygen consumption during isometric contractions of the knee extensors determined with near-infrared spectroscopy. J Appl Physiol. 2005;99:579-586.
Dankel SJ, Mattocks KT, Jessee MB, Buckner SL, Mouser JG, Loenneke JP. Do metabolites that are produced during resistance exercise enhance muscle hypertrophy? Eur J Appl Physiol. 2017;117(11):2125-2135.
Meyers CR. Effects of two isometric routines on strength, size, and endurance in exercised and nonexercised arms. Res Q Exerc Sport. 1967;38(3):430-440
Balshaw TG, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specific functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol. 2016;120(11):1364-1373.
Schott J, McCully K, Rutherford OM. The role of metabolites in strength training: short versus long isometric contractions. Eur J Appl Physiol Occup Physiol. 1995;71(4):337-341.
Kongsgaard M, Aagaard P, Kjaer M, Magnusson SP. Structural Achilles tendon properties in athletes subjected to different exercise modes and in Achilles tendon rupture patients. J Appl Physiol (1985). 2005 Nov;99(5):1965-71. doi: 10.1152/japplphysiol.00384.2005. Epub 2005 Aug 4. PMID: 16081623.
Arampatzis A, Karamanidis K, Albracht K. Adaptational responses of the human Achilles tendon by modulation of the applied cyclic strain magnitude. J Exp Biol. 2007;210:2743-2753.
Arampatzis A, Peper A, Bierbaum S, Albracht K. Plasticity of human Achilles tendon mechanical and morphological properties in response to cyclic strain. J Biomech. 2010;43(16):3073-3079.
Burgess KE, Connik MJ, Graham-Smith P, Pearson SJ. Plyometric vs isometric training influences on tendon propertied and muscle output. J Strength Cond Res. 2007;21(3):986-989.
Kubo K, Kanehisa H, Fukunaga T. Effects of different duration isometric contractions on tendon elasticity in human quadriceps muscles. J Physiol. 2001;536(2):649-655.
Kubo K, Ishigaki T, Ikebukuro T. Effects of plyometric and isometric training on muscle and tendon stiffness in vivo. Physiol Rep. 2017;5(e13374):1-13
Massey G, Balshaw T, Maden-Wilkinson T, Tillin N, Folland J. Tendinous tissue adaptation to explosive- vs. sustained-contraction strength training. Front Physiol. 2018;9(1170):1–17.
Bandy WD, Hanten WP. Changes in torque and electromyographic activity of the quadriceps femoris muscles following isometric training. Phys Ther. 1993;73(7):455-465.
Barak Y, Ayalon M, Dvir Z. Transferability of strength gains from limited to full range of motion. Med Sci Sports Exerc. 2004;36(8):1413-1420.
Massey CD, Vincent J, Maneval M, Moore M, Johnson JT. An analysis of full range of motion vs. partial range of motion training in the development of strength in untrained men. J Strength Cond Res. 2004;18(3):518-521.
Tillin NA, Folland JP. Maximal and explosive strength training elicit distinct neuromuscular adaptations, specific to the training stimulus. Eur J Appl Physiol. 2014;114(2):365-374.
Maffiuletti NA, Martin A. Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Med Sci Sports Exerc. 2001;33(7):1220-1227
Alegre LM, Ferri-Morales A, Rodriguez-Casares R, Aguado X. Effects of isometric training on the knee extensor moment– angle relationship and vastus lateralis muscle architecture. Eur J Appl Physiol. 2014;114(11):2437-2446.
Bogdanis GC, Tsoukos A, Methenitis SK, Selima E, Veligekas P, Terzis G. Effects of low volume isometric leg press complex training at two knee angles on force-angle relationship and rate of force development. Eur J Sport Sci. 2018;1-9. https://doi.org /10.1080/17461391.2018.1510989. [Epub ahead of print].
Behm DG, Sale DG. Intended rather than actual movement velocity determines velocity-specific training response. J Appl Physiol. 1993;74(1):359-368.
Balshaw T, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specific functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol (1985) 2016; 120: 1364–1373
Davies J, Parker DF, Rutherford OM, Jones DA. Changes in strengh and cross sectional area of the elbow flexors as a result of isometric strength training. Eur J Appl Physiol 1988; 57: 667–670
Garfinkel S, Cafarelli E. Relative changes in maximal force, EMG, and muscle cross-sectional area after isometric training. Med Sci Sports Exerc 1992; 24: 1220–1227
Ikai M, Fukunaga T. A study on training effect on strength per unit corss-sectional area of muscle by means of ultrasonic measurement. Eur J Appl Physiol 1970; 28: 173–180
Jones DA, Rutherford OM. Human muscle strength training: The effects of three different regimes and the nature of the resultant changes. J Physiol 1987; 391: 1–11
Kanehisa H, Nagareda H, Kawakami Y, Akima H, Masani K, Kouzaki M, Fukunaga T. Effects of equivolume isometric training programs comprising medium or high resistance on muscle size and strength. Eur J Appl Physiol 2002; 87: 112–119
Kubo K, Ohgo K, Takeshi R, Yoshinaga K, Tsunoda N, Kanehisa H, Fukunaga T. Effects of isometric training at different knee angles on the muscle-tendon complex in vivo. Scand J Med Sci Sports 2006; 16: 159–167
Noorkoiv M, Nosaka K, Blazevich AJ. Neuromuscular adaptations associated with knee joint angle-specific force change. Med Sci Sports Exerc 2014; 46: 1525–1537
Noorkoiv M, Nosaka K, Blazevich AJ. Effects of isometric quadriceps strength training at different muscle lengths on dynamic torque production. J Sports Sci 2015; 33: 1952–1961
Schott J, McCully K, Rutherford OM. The role of metabolites in strength training II. Short vs. long isometric contractions. Eur J Appl Physiol 1995; 71: 337–341
Abe, Takashi, Kumagai, Kenya, Brechue, William F. Fascicle length of leg muscles is greater in sprinters than distance runners, Medicine & Science in Sports & Exercise: June 2000; 32(6): 1125-1129.
Kumagai K, Abe T, Brechue WF, Ryushi T, Takano S, Mizuno M. Sprint performance is related to muscle fascicle length in male 100-m sprinters. J Appl Physiol (1985). 2000 Mar;88(3):811-6. doi: 10.1152/jappl.2000.88.3.811. PMID: 10710372.
Lum, D., Barbosa, T.M., Joseph, R. et al. Effects of Two Isometric Strength Training Methods on Jump and Sprint Performances: A Randomized Controlled Trial. J. of SCI. INSPORTANDEXERCISE3, 115–124 (2021). https://doi.org/10.1007/s42978-020-00095-w
Balshaw T, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specifc functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol. 2016;120(11):1364–73
Tillin NA, Folland JP. Maximal and explosive strength training elicit distinct neuromuscular adaptations, specifc to the training stimulus. Eur J Appl Physiol. 2014;114(2):365–74.
Ronnestad BR, Kvamme NH, Sunde A, Raastad T. Short-term efects of strength and plyometric training on sprint and jump performance in professional soccer players. J Strength Cond Res. 2008;22(3):773–80
Behm DG, Sale DG (1993) Intended rather than actual movement velocity determines velocity-specific training response. Journal of Applied Physiology 74(1):359–68.
Maffiuletti NA, Martin A (2001) Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Medicine and Science in Sports and Exercise 33(7):1220–27.
Olsen PD, Hopkins WG (2003) The effect of attempted ballistic training on the force and speed of movements. Journal of Strength and Conditioning Research 17(2):291–98.
Viitasalo JT, Komi PV (1981) Effects of fatigue on isometric force- and relaxation-time characteristics in human muscle. Acta Physiologica Scandavica 111(1):87–95.
Oranchuk DJ, Storey AG, Nelson AR, Cronin JB. Isometric training and long-term adaptations: Effects of muscle length, intensity, and intent: A systematic review. Scand J Med Sci Sports. 2019 Apr;29(4):484-503. doi: 10.1111/sms.13375. Epub 2019 Jan 13. PMID: 30580468.
Olen pääkaupunkiseudulla vaikuttava voima-, fysiikka- ja yleisurheiluvalmentaja. Valmentajana olen toiminut kohta 10 vuoden ajan. Olen erityisen kiinnostunut urheilijoiden suorituskyvystä ja harjoittelun yksilöllisistä eroista urheilijoilla sekä periodisaatiosta. Saan urheilijat suoriutumaan lajissaan paremmin.
Tämä sivusto on luotu aputyökaluksi valmentajille, jotka haluavat perustaa valmentamisensa tieteelliseen näyttöön. Alalla on vielä paljon käytössä vanhentuneita toimintatapoja, jotka heikentävät urheilijoiden suorituskykyä. Pyrin jakamaan tieteellistä tietoa ja auttamaan objektiivisesti kaikkia valmentajia.