Nopeusperustainen harjoittelu, eli Velocity-based training (VBT), on viime aikoina tullut erittäin suosituksi. Tangon liikenopeuden mittaamiselle on useita eri käyttötarkoituksia, kuten esimerkiksi nopeuden monitorointi harjoittelun aikana, oikean liikenopeusspesifin adaptaation rakentaminen, urheilijan testaaminen tai sarjojen tekeminen tiettyyn nopeuden vähenemiseen saakka.
Uusia laitteita on tullut runsaasti markkinoilla ja niiden hinnat ovat tulleet huomattavasti alaspäin viime vuosina. Mittausvälineissä on kuitenkin paljon eroja ja jotkin laitteet ovat aivan turhia. Mihin valmentajan kannattaa sijoittaa rahansa, jotta rahoille saa mahdollisimman paljon vastinetta?
Kiihtyvyysanturitekniikkaan perustuvat laitteet näyttävät hyviltä ja toimivilta, mutta ovat epätarkkoja.
Lyhyt vastaus:
Älä tuhlaa rahojasi kiihtyvyysantureihin vaan sijoita lineaarisen asennon sijaintiantureihin. Esimerkiksi: GymAware, Tendo Unit tai T-force ovat hyviä vaihtoehtoja.
Fysiikkavalmennus.fi suosittelee:
GymAware – laitetta.
+ Kestävä, todettu validiksi, hyvä applikaatio.
- Kallis, sovellukset vain mac:ille.
GymAware on pieni ja sitä on helppo kuljettaa mukana.
Musta hevonen:
Puhelinapplikaatiot. Erittäin edullisia ja helppokäyttöisiä. Monet ovat vielä epätarkkoja, mutta päivitysten myötä tarkkuus voi mahdollisesti parantua? Esimerkiksi MyLift:in saa 15 dollarin sijoituksella itselleen. Toisaalta sovelluksissa on vielä todella isoja virhemarginaaleja, mutta edullisuus tekee niistä houkuttavia.
VBT - mittautekniikoiden esittely ja kattava laitelista sekä pidemmät perustelut VBT-laitteiden tarkkuudelle.
Kultainen standardi – 3D kuvaaminen
Tarkin tapa mitata liikenopeutta on liikkeen kuvaaminen (5,6). Kuvaaminen mittaa myös liikkeessä tapahtuvat rotaatiot ja muut kierrot, mitä mitkään muut menetelmät eivät mittaa. Kuvaamisella on mahdollista saada lisää tietoa liikkeen laadusta. Kamerajärjestelmän kanssa voi prosessoida nivelkulmia, tangon liikenopeutta ym. Esimerkkinä tällaisesta järjestelmästä on Vicon – järjestelmä. Täydellisessä maailmassa urheilijat tekisivät voimalevyn päällä suorituksen, ja valmentajalla olisi kymmenen Vicon kameraa ja järjestelmä pyörittämässä analyysiä. Tällaisen hinta on karkeasti noin 40 000 – 60 000 euroa, joten luultavasti tuohon ei ainakaan minun budjetilla ihan heti päästä. Noin kymppitonnilla pääsee liikkeelle Vicon:in kanssa, mutta käytännössä puhutaan siis laboratoriokäytöstä. Esimerkiksi Jyväskylän Yliopistossa Lahti et al. (2018) ovat tutkineet takakyykyn kinematiikkaa Vicon-järjestelmällä.
Kuvaamisen plussat: tarkkaa dataa ja mahdollisuus lisätä analyysiin liikkeet rotaatiot. Harva liike liikkuu suoraan ylös- tai alaspäin, joten virhemarginaalia muodostuu kaikissa muissa mittaustavoissa, jos mittauksia ei tehdä smith-laitteessa.
Haittoja tuossa on muutamia. Esimerkiksi menetelmän tarkkuus aiheuttaa myös ongelmia. Menetelmä mittaa tangon siirtymää myös horisontaalitasossa. Tämä vaikuttaa nopeuden mittaamiseen ja voi tehdä siitä haastavaa. Analyysia ei voi tehdä mittaustilanteen kanssa samaa aikaan ja palaute tulee jälkikäteen. Valmentaja ja urheilija tarvitsee yleensä palautteen välittömästi, joten 3D kuvaaminen ei ole siis käytännöllinen valmennustilanteeseen. Lisäksi laitteet prosessoivat hyvin paljon dataa ja tämän takia alla tulee olla erittäin suorituskykyinen tietokone.
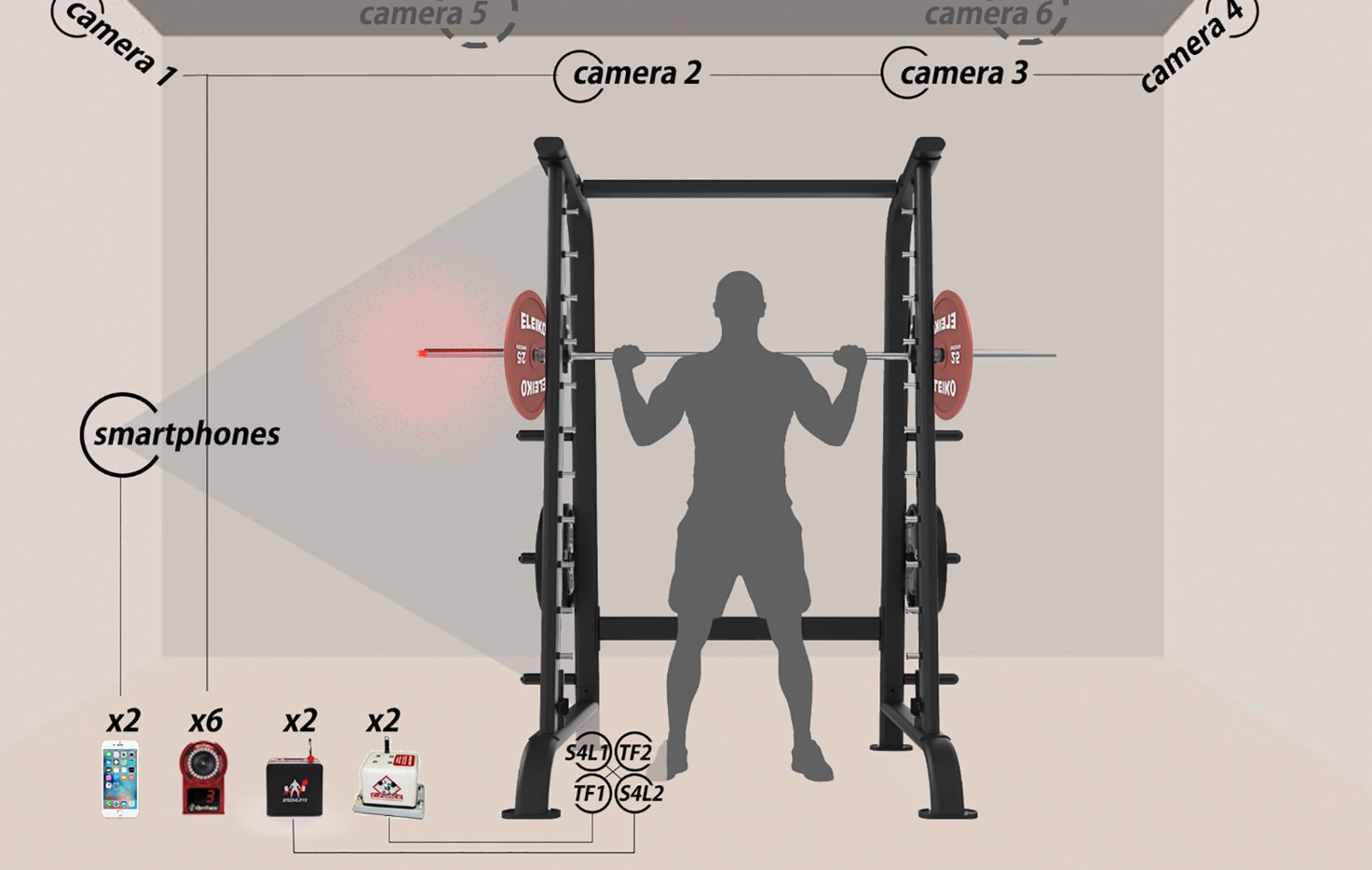
Tutkimusasetelma siitä, miten mitataan tangon liikenopeutta erilaisilla tekniikoilla. Tässä asetelmassa oli mukana kuusi kameraa, kaksi puhelinsovellusta, kaksi T-Forcec mittaria ja kaksi Speed4Lift mittaria.
Lineaarisen asennon ja nopeuden sijaintianturi
Linear position transducers (LPT) tai linear velocity transducers (LVT), eli vapaasti suomennettuna lineaarisen asennon ja nopeuden sijaintianturit ovat valmentajalle tällä hetkellä ehdottomasti paras valinta. Näitä pidetään ”alkuperäisinä” VBT laitteina. Niitä on ollut olemassa vuosikymmenen ajan niiden yksinkertaisuuden, käyttäjälähtöisyyden ja datan nopean prosessoinnin takia. Tulokset ovat välittömästä nähtävissä. Käytännössä nämä toimivat niin, että naru kiinnitetään esimerkiksi tankoon ja toinen pää mittauslaitteeseen. Kun tankoa liikutetaan niin narukin liikkuu, joka liikuttaa mittauslaitteessa olevaa kelaa. Kelan pyörimisen nopeutta mittaa enkooderi, joka mittaa kelan rotationaalista siirtymää ja aikaa. Tämän perusteella pystytään hyvin simppelisti määrittämään lineaarinen nopeus. Haittapuolia laitteissa on, etteivät ne mittaa horisontaalista siirtymää. Lähes kaikissa liikkeissä on horisontaalista liikettä ja harva liike kulkee täysin pystysuoraan. Jos haluaa todella tarkkoja tuloksia niin sitten kannattaa käyttää pystysuoraa smith-laitetta, jossa pystyy tekemään tangolla vain vertikaalista liikettä. Toisaalta pieni heilunta vapaalla tangolla haittaa mittauksen tarkkuutta vain hiukan, joten käytännön valmennuksessa sillä ei ole juurikaan vaikutusta. Mittauslaitteiston hyötyjä on laitteen yksinkertaisuus, helppokäyttöisyys, tarkkuus ja datan välitön saatavuus. Lisäksi sovellukset ovat nykyään helppokäyttöisiä ja käytännönläheisiä sekä laitteiden hinnat ovat tulleet viime vuosina runsaasti alaspäin.
Kiihtyvyysantureita löytyy nykypäivänä monesta eri elektronisista laitteista. Esimerkiksi kännykkä laskee askeltesi määrän kiihtyvyysanturilla. Näitä samoja siruja myydään nyt erillisissä pakkauksissa, joita voi kiinnittää tankoon. Puhelimen ja sirun kanssa voi mitata tangon tai urheilijan liikenopeutta. Kiihtyvyyssmittareissa on valtavan suuria virhemarginaaleja ja niitä ei voi millään tavalla suositella harjoittelun monitoroimiseksi. Esimerkiksi yksi tutkimus arvioi (3) kiihtyvyysanturit (Push:in ja Beast:in) kaikista epätarkimmiksi tavoiksi mitata tangon liikenopeutta. Saman tutkimuksen mukaan LPT ja LVT, kamerakuvausjärjestelmät sekä puhelinapplikaatot olivat tarkkoja nopeuden mittaamistapoja rajoitetussa lineaarisessa liikkeessä (smith laitteessa). Kiihtyvyysanturit todettiin tutkimuksessa epätarkoiksi ja niitä ei voi suositella. Myös muut (7) ovat todenneet kiihtyvyysanturit erittäin epätarkoiksi varsinkin kevyemmillä kuormilla.
Puhelinapplikaatiot
Lähes jokaisen taskusta löytyy nykyään älypuhelin, joka mahdollistaa myös tangon liikenopeuden mittaamisen kameran avulla. Applikaatiot, jotka mahdollistavat tämän ovat edullisia ja tarkkuudeltaan kiihtyvyysantureiden vertaisia. MyLift on esimerkki tällaisesta sovelluksesta. Sovelluksissa on osoitettu olevan vielä todella isoja virhemarginaaleja. Esimerkiksi MyLift sovelluksen virhemarginaali oli pahimmillaan > 0.10 m/s, SDC > 0.23 m/s (1). Tarkoittaen sitä, että heitto voi olla ensimmäisessä mittauksessa noin 10 prosenttia alaspäin ja toisessa mittauksessa 10 prosenttia ylöspäin. Tämä tarkoittaisi 20 prosentin kehitystä tietyssä ajassa, vaikka todellisuudessa urheilija olisi täysin samalla tasolla. Tällaiset virhemarginaalit mittauslaitteissa voivat johtaa virhearviointeihin harjoittelun suunnittelussa ja pahimmillaan ohjata toimintaa ihan väärään suuntaan. Toisaalta sovelluksen aikaisempi versio PowerLift on osoitettu olevan hyvinkin tarkka ja virhemarginaali on aika minimaalinen (0.008 ± 0.03 m · s−1) (4). Toisaalta tutkimuksessa vertailtiin applikaatiota vain yhteen LPT laitteeseen. Tarkemmat tulokset saisi, kun vertailtaisiin useaan eri mittauslaitteeseen. Ja isona huomiona, että tutkimuksen teki sovelluksen kehittäjä, joka ei ihan vastaa kaikkia eettisiä periaatteita. Hyödyt: todella halpoja. Haitat: erittäin epätarkkoja.
Taulukko 1: Yhteenvedot eri mittausmenetelmistä.
Hyödyt
Haitat
Esimerkkituotteet
3D kuvaaminen
Tieteellisesti tarkin.
Kallis ja ei käytännöllinen. Laboratioriokäyttöön.
Vicon, Dartfish
2D kuvaaminen
Halpa ja helppo.
Ei tarkka ja sovelluksia vielä todella vähän.
RepOne, GymAware, Tendo unit, T-Force, Musclelab, Speed4lifts, Chronojump.
Kiihtyvyysanturit
Erittäin edullisia.
Epätarkkoja.
Beast, Push, Barsensei
Fysiikkavalmennus.fi suosittelee GymAwarea:
Gymaware on todettu tieteellisen tarkaksi ja vain alle 40% 1RM kuormista saattaa tulla pientä heittoa (2). Lisäksi GymAwawren sovellus on todella hyvä ja käyttöliittymä vaivaton. Esimerkiksi joukkueelle saa helposti käyttöön leader boardin, jossa näkee kuka tuottaa parhaimman nopeuden. Iso miinus siitä, että ohjelmisto pelkästään Applelle. Myös muut LPT laitteet tarkkoja, esimerkiksi T-force, Tendo ja Musclelab (1, 7). Uusista laitteista erittäin lupaavia ovat RepOne ja Speed4Lifts. Kummatkin maksavat alle 400 euroa ja vaikuttavat olevan valideja laitteita. Odotetaan vielä tutkimustuloksia uusista halvemman hintaluokan laitteista. Kaikki kiihtyvyysanturin ovat aikalailla kuraa ja niitä ei kannata ostaa.
Miksi itse olen ostanut Tendon? Gymaware olisi paras, mutta en käytä Applea niin olis pitänyt ostaa tabletit ja muut vastaavat vekottimet vielä laitteen lisäksi. Lisäksi en tykkää siitä, että pitää maksaa vuosimaksu, jos haluaa hallita useampia joukkueita. Tendo on tosi edullinen laatunsa nähden. Itse ostin Bluetooth painonnostoversion joka oli muistaakseni 1250 euroa. 1000 eurolla olisi saanut laitteen ilman pudotussuojaa. Laite antaa tarkasti ja nopeasti tarvittavan datan. Haittoja laitteessa on sovelluksen yksinkertaisuus, joka ei mahdollista hirveästi temppuja. Lisäksi Tendo on aika iso ja painava, minkä olen huomannut vaikuttavan käyttökokemukseen. Nyt ostaisin pienemmän, jota jaksaisi aina kantaa mukana pitkin kyliä.
Musta hevonen:
Puhelinapplikaatiot. Edullisia ja helppoja. MyLift data ristiriitaista ja luultavasti sovelluksessa vielä isoja virhemarginaalejakin (1,4). Ja pakko vielä lisätä, että toisen tutkimuksen, missä sovellus osoitettiin tarkaksi, teki sovelluksen kehittäjä, joten tuloksia voi hiukan kyseenalaistaakin. Muita heikkouksia tuossa on, että vaatii urheilijan liikelaajuuden mittaamista ennen kuvaamista. Täältä sovelluksen saa esimerkiksi noin 15 dollarilla. Puhelinten kamerat ja erilaiset tracking-ominaisuudet mahdollistaisivat jo nyt laadukkaan ja tarkan sovelluksen luomista, joten odotetaan uusia avauksia tulevaisuudessa.
Vielä lopuksi listattuna suurin osa tällä hetkellä markkinoilla olevista laitteista:
Taulukko 2: Lähes kaikki LPT ja kiihtyvyysanturitekniikkaa hyödyntävät laitteet listattuna. Tästä listasta on helppo valita itselle suosikki. Hintoihin tulee usein päälle vielä kuljetusmaksut.
+ Käytetty runsaasti eri tutkimuksissa välineenä. Lue lisää täältä. Taitaa olla ainoa laite, joka ennustaa 1RM:n suhteellisen tarkasti submaksimaalisesta kuormasta. Näyttää myös välittömän nopeuden vähentymisen edelliseen sarjaan verrattuna. - nettisivut aika alkeelliset (katso itse :D), en ole varma onko ohjelmistoa englanniksi
Osaatko koodailla? Jos osaat niin osta ehdottomasti tämä. Tarkka ja erittäin hyvä laite. Chronojumpin kaikki laitteet perustuvat samalle idealle, että laitteet ovat hyviä ja edullisia, mutta valmista ohjelmistoa ei ole. Eli itse pitää vähän osata koodailla. Raakaa dataa tulee kyllä hyvin.
+ Sovelluksessa laajat ominaisuudet; mm. tangon liikeradan merkintä. Saa tankoon kiinni. Painojen vaihto helppoa. - Sovellus vain Apple tuotteille. Valmentajalle oma sovellus, joka on maksullinen (30 dollaria kuukaudessa).
Lähteet:
Martínez-Cava, A., Hernández-Belmonte, A., Courel-Ibáñez, J., Morán-Navarro, R., González-Badillo, J. J., & Pallarés, J. G. (2020). Reliability of technologies to measure the barbell velocity: Implications for monitoring resistance training. PloS one, 15(6), e0232465. https://doi.org/10.1371/journal.pone.0232465
Orange, S., Metcalfe, J., Marshall, P., Vince, R., Madden, L. & Liefeith, A. (2018). Test-Retest Reliability of a Commercial Linear Position Transducer (GymAware PowerTool) to Measure Velocity and Power in the Back Squat and Bench Press. Journal of Strength and Conditioning Research. 34. 1. 10.1519/JSC.0000000000002715.
Pérez Castilla, A., Piepoli, A., Delgado, G. & Garrido, G. & García, R. (2019). Reliability and Concurrent Validity of Seven Commercially Available Devices for the Assessment of Movement Velocity at Different Intensities During the Bench Press. The Journal of Strength and Conditioning Research. 33. 10.1519/JSC.0000000000003118.
Balsalobre-Fernández, C., Marchante, D., Muñoz López, M. & Saiz, S. (2017). Validity and reliability of a novel iPhone app for the measurement of barbell velocity and 1-RM on the bench-press exercise. Journal of Sports Sciences. 36. 10.1080/02640414.2017.1280610.
Lorenzetti, S., Lamparter, T. & Luthy F. (2017) Validity and reliability of simple ¨ measurement device to assess the velocity of the barbell during squats. BMC Res Notes 10: 707.
Weakley. J., Wilson, K., Till, K., Read, D., Darrall-Jones, J., Roe, G., et al. (2017) Visual feedback attenuates mean concentric barbell velocity loss, and improves motivation, competitiveness, and perceived workload in male adolescent athletes. J Strength Cond Res. Epub ahead of print.
Banyard, H., Nosaka, K., Sato, K. & Haff, G. (20179 Validity of various methods for determining velocity, force, and power in the back squat. Int J Sports Physiol Perform 12: 1170–1176.
Viime osassa syvennyttiin pikajuoksun voimaharjoittelun teoriaan. Nostona huomio siitä, ettei korkean voiman ja suuren liikenopeuden periodisointi peräkkäin ole järkevää pikajuoksussa, vaan niitä kannattaisi kehittää rinnakkain kokoajan. Tämä johtuu pääasiassa siitä, että eri lihakset tarvitsevat erilaista voimantuottokykyä. Tässä osassa keskitytään käytännön puoleen ja siihen, miten edellä mainittuja asioita voi hyödyntää käytännössä sekä miten voimaharjoitteluohjelma kannattaisi rakentaa pikajuoksijalle. Tiedossa on paljon videomateriaalia ja käytännön esimerkkejä.
Alustus: mitä treenataan ja miksi?
Pikajuoksija ei välttämättä tarvitse kyykkyä ollenkaan
Kyykkyä pidetään hyvänä liikkeen pikajuoksijalle. Sen kanssa kannattaa edetä kuitenkin varovasti. Kyykky on hyvä perusvoimapohjien rakentaja, mutta esimerkiksi hypertrofiassa kehittää lähes täysin etureisien distaalisia päitä. Kanesha et al. (2003) seurasivat juniori painonnostajia 18 kuukauden ajan ja huomasivat, että reiden ojentajien poikkipinta-ala kasvoi merkitsevästi distaalisista päistä mittauksissa, mutta ei proksimaalisissa. Painonnostajat kyykkäilivät syvää high-bar kyykkyjä (kuva 1). Pikajuoksija ei tarvitsisi distaaliseen päähän kokoa vaan nimenomaan proksimaaliseen päähän. Huippujuoksijoiden morfologiaa vertaillessa huonompi tasoisiin juoksijoihin voidaan huomata, että paremmilla juoksijoilla (10–10.9s) on suurempi lihastiheys yläosassa reittä, eikä merkitseviä eroja vastus lateraliksen paksuudessa, eli distaalisessa päässä (Kumagai et al., 2010). Lisäksi paremmilla juoksijoilla on huomattu negatiivinen yhteys lihaksen fasiculuksen pituuden ja ennätysajan välillä, tarkoittaen sitä, että mitä pidempi lihassäie on sitä parempi ennätysaika on. Lihaspaksuus ei siis välttämättä tässä auta nopeuden kehittämisessä.
Kuva 1: Runsas kyykkyharjoittelu ei aiheutta pikajuoksijalle toivottuja adaptaatioita. Syvä high-bar kyykky aiheuttaa erityisesti reiden distaalisen pään kasvua, mutta ei proksimaalista. Eri kyykkyvariaatioilla voidaan saada erilaisia adaptaatioita eri kohtaan lihasta aikaan.
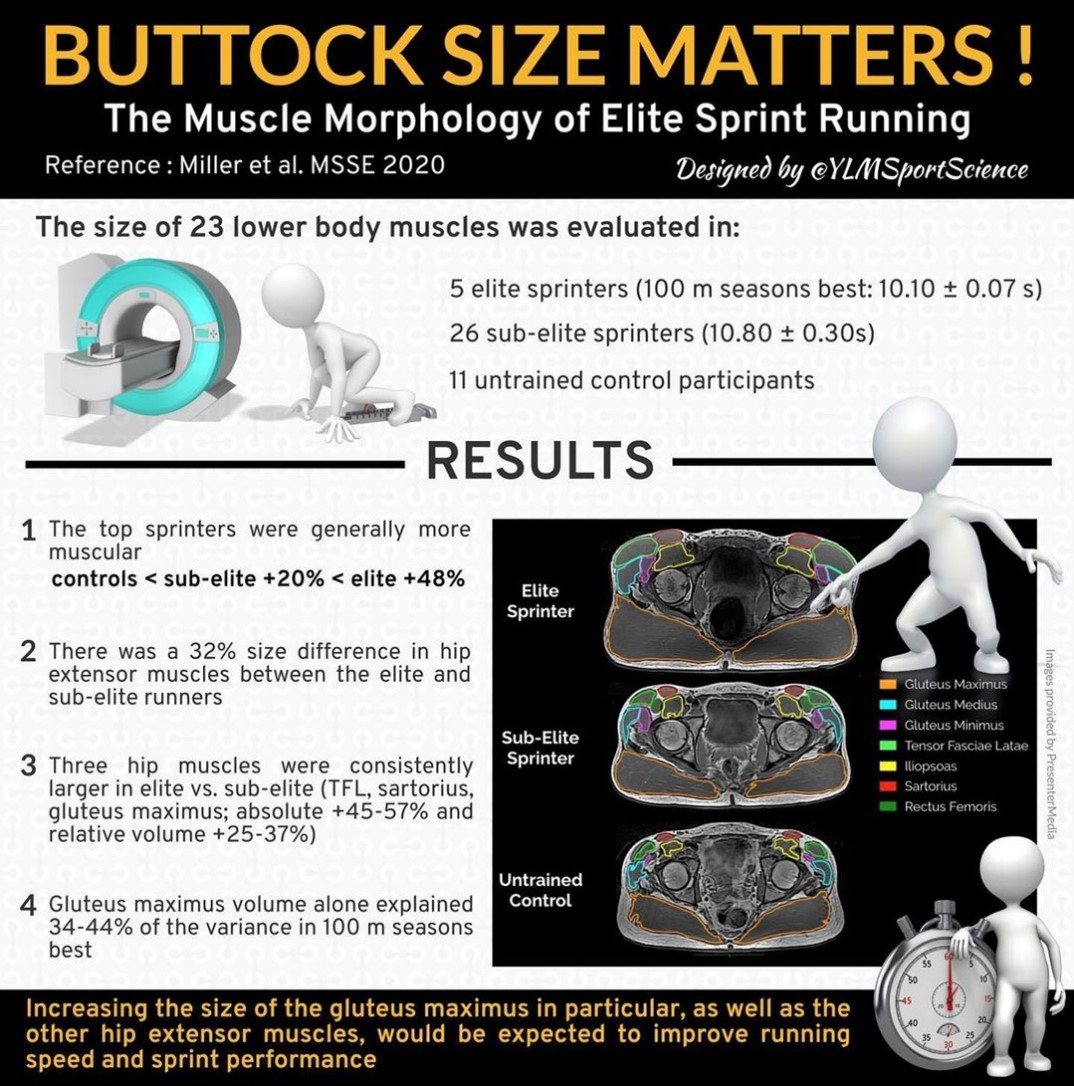
Pikajuoksijan moottori on pakara
Pikajuoksijan tehontuotto välineen toimii pakara. Mitä nopeampi juoksija on, sitä isompi pakara häneltä löytyy (kuva 2). Tästä voidaan päätellä, että isompaa pakaraa tarvitaan nopeampaan juoksuun. Lisäksi kovempi tasoisilla juoksijoilla (10,10 ± 0,07) on selvästi pienempi rasvaprosentti verrattuna keskitason pikajuoksijoihin (10,80 ± 0,30). Rasvaprosentin alentaminen onkin yksi helpoin tapa parantaa juoksunopetta. Liikuteltavan massan määrä pienenee, mutta tehontuotto pysyy samana.
Kuva 2: YLMSportSciencen infograafi Millerin ja kumppanien (2020) tutkimuksesta, jossa havaittiin, että huippupikajuoksijoilla verrattuna kansallisen tason sprinttereihin on enemmän lihasmassaa, isommat pakaralihakset ja pienempi rasvaprosentti.
Pakaraa voi kehittää lantionnostoilla ja sen eri variaatioilla. Tässä esimerkkinä pikajuoksija Annimari Korte ja 255 kilon lantionnosto.
Vastuskelkkaharjoittelu on hyvä tapa kehittää lajivoimaa. Usein vastuskelkkaharjoittelussa käytetään noin 10-15% painoja kehonpainosta. Tavoitteena on yleensä, että maksiminopeus ei tippuisi enempää kuin 10%. Toisaalta horisontaaliseen voimaharjoitteluun erittäin raskas kelkkaharjoittelu voisi olla tehokas väline. Horisontaalista voimaharjoittelua on hankalaa toteuttaa salilla. Raskas vastuskelkkaharjoittelu mahdollistaa tilanteen, jossa voi kehittää suuria voimia haluttuun kulmaan (eteenpäin kallistuneeseen) ja saada aikaan suuren lihasaktiivisuuden alaraajojen lihaksistossa. Kevyt vastuskelkkaharjoittelu ei mahdollista suuren voiman tuottamista ja ei kehitä voimantuottokykyä. Raskas vastuskelkkaharjoittelu (80% kehonmassasta kelkassa) on todettu olevan tehokas keino kehittämään maksimaalista horisontaalista voimantuottokykyä (Morin et al., 2016). Toivoisinkin näkeväni enemmän myös raskasta kelkkaharjoittelua pelkän kevyeiden kelkkajuoksujen rinnalla.
Raskaat kelkkavedot, jossa kuorma on oikeasti raskas, voimantuottoaika korkeampi ja pystytään kehittämään voimantuottokykyä horisontaalisesti.
Kevyet kelkkajuoksut taas toimivat enemmän lajivoiman ja nopeuden kehittämisessä, eivät niinkään voimantuottokyvyn kehittämisessä.
Vastuskelkkaharjoittelu saattaa olla tehokkaampaa horisontaalisen voiman ja tehontuoton kehittämistä kuin perinteinen voimaharjoittelu (Petrakos ym., 2016). Tämä johtuisi pääasiassa siitä, että vastustkelkkaharjoittelu toistaisi samaa motorista kaavaa ja lihassolujen supistustyyppiä kuin pikajuoksu. Vastuskelkkaharjoittelu jaetaan yleensä kevyeeseeen (< 10% nopeuden väheneminen), keskikovaan (< 10-15%), raskaaseen (< 15-30%) ja todella raskaaseen (> 30%) kuormaan (Petrakos et al., 2016). Osan perinteistä voimaharjoittelusta voi korvata raskaalla vastuskelkkavedoilla (Cross et al., 2018). Cross ja kumppanit (2018) ehdottavat kuormaksi sellaista, että vauhti putoaa noin 50% vähennystä maksiminopeudessa. Morin ja kumppanit (2017) testasivat tätä käytännössä ja huomasivat selvän eron horisontaalisessa voimantuottokyvyssä, kun verrattiin normaalia juoksuja tehneisiin kontrolliryhmään. Mielenkiintoista olisi ollut nähdä vertaus perinteistä voimaharjoittelua tehneeseen ryhmään.
Taulukko 1: Vastuskelkkajuoksut (resisted sprints) vaativat 3-6 minuutin tauon suoritusten välissä ja vähintään pari päivää harjoitussessioiden välissä. Kokonaisvoluumin on syytä olla matala yhdellä harjoituskerralla noin 50-200 metriä. Haugen ym., 2019.
Training method
Distance (m)
Intensity (%)
Recoveries (min)
Total session volume (m)
Initiation
Time to next HIS (hours)
Footwear and surface
Acceleration
10–50
> 98
2–7
100–300
Block/3-point/crouched
48
Spikes on track
Maximal velocity
10–30a
> 98
4–15
50–150a
20–40-m flying start
48–72
Spikes on track
Sprint-specific endurance
80–150
> 95
8–30
300–900
Standing start
48–72
Spikes on track
Speed endurance
60–80
90–95
2–4 (8–15)
600–2000
Standing start
48–72
Spikes on track
Resisted sprints
10–30
80–95b
3–6
50–200
3-point/crouched
48
Optional
Assisted sprints
10–30a
≤ 105
5–15
≤ 100a
20–40-m flying start
48
Spikes on track
Tempo
100–300
60–70
1–3
1000–2000
Standing start
24
Trainers on grass
a. Intensity is expressed in percent of maximal velocity. Recovery = time between repetitions (sets). HIS = high-intensive session b. Flying start distance excluded c. The perceived effort is maximal, so the velocity decline is caused by resistance loading
Voimaharjoittelua lonkan ojentajille ja koukistajille korkeammilla nopeuksilla
Horisontaalivoimaa ja lonkan aluetta tulee kehittää erityisesti myös nopeammilla liikenopeuksilla kuten ensimmäisessä osassa käytiin läpi. Toisin kuin tulevissa videoissa niin jokainen liike tulee suorittaa maksimaalisella liikenopeudella!
Kuminauha kahvakuulaheilautus:
Lonkan koukistus – voi tehdä myös ojennuksen – mallintaa heilausvaihetta (kevyt kuorma, nopeasti!):
Näitä kannattaa tehdä harjoittelun loppupuolella kun on ensiksi tehty nopean liikenopeuden liikkeet. Polven alueen lihaksisto ottaa juoksun aikana suuria voimia vastaan eksentrisesti ja niiden vahvistaminen on äärimmäisen tärkeää.
Lisätty eksentrinen kuorma takareisikoukistukseen. Kannattaa käyttää sellaista painoa, jota yhdellä jalalla ei saisi ylös:
Reverse nordic, eli etureiden harjoittamista eksentrisesti. Haastetta saa kasaamalla lisäpainoa syliin.
Isometriset takareisille. Näihin kannattaa rakentaa progressio niin, että tavoitteena on käyttää lisäpainona 50% kehonpainosta ja tavoitteena pitää helposti yhdellä jalalla 30s asentoa yllä. Tarkoittaen, että 100 kiloinen mies jaksaisi pitää helposti 50 kilon kuormaa ja 30s asentoa yllä yhdellä jalalla.
Vertikaalivoima
Vertikaalivoimaa on järkevä kehittää sopivissa määrin. Erityisesti pakaran ja pohkeiden harjoittaminen on tärkeää pikajuoksun kannalta.
Askelkyykky korokkeelta ja lisäpainoa rohkeasti niskaan. Pikajuoksija voi vielä progressoida liikettä lähtemällä polvennostoasennosta ja kaatumalla eteenpäin, jotta maksimoidaan eksentrinen kuormitus:
Romanialainen maastaveto (voimantuottokäyrää voi muokata kuminauhoilla tai ketjuilla) on erinomainen like takaketjun kehittämiseen. Tästä puoli unilateraalinen versio, eli split stance Romanian deadlift. Tässä toinen jalka antaa hieman tukea. Tykkään tosi paljon itse laittaa tätä urheilijoille. Asento mahdollisaa suuremman painon käyttämisen verrattuna yhden jalan versioon, koska tasapaino pysyy paremmin, mutta silti pystyy keskittymään yhden jalan tekemiseen paremmin kuin kahdella jalalla.
Kuntopallovariaatioita on monia, mutta tässä yksi. Kuntopallon heitto ylöspäin. Näissä vain mielikuvitus on rajana.
Tempaus työntöotteella roikunnasta:
Raaka rinnalleveto roikunnasta:
Boksille nousu hypyllä. Lisäpainoa taas käsipainoista.
Trap-bar hypyt:
Tankohypyt puolikyykystä:
Työntöveto:
Flywheel laitteilla takareidet. Laitteiden käyttö perustuu nopeammalle ja aggressiivisemmalle eksentriselle vaiheella, mikä auttaa pikajuoksijaa kehittämään nimenomaan jarrutavia voimantuotto ominaisuuksia, mitä tarvitaan polven alueen lihaksistolle. Mitä enemmän ja nopeammin tuottaa voimaa konsentrisessa vaiheessa, sitä enemmän kiekko pyörii, ja vetää taas urheilijaa eksentrisessä vaiheessa alaspäin. Täten eksentrinen vaihe on nopeampi ja vaativampi verrattuna normaaliin voimaharjoitteluun. Eksentrinen vaihe saakin aikaan vauhtipyöräharjoittelun pääharjoitusvasteen.
Pohkeiden harjoitteleminen monipuolisesti. Esimerkiksi erilaisten juoksudrillien tekeminen lisäpainon kanssa joko niskassa tai pään päällä. Lisäksi isometrinen harjoittelu on erittäin tehokasta pohkeille:
Tässä oli listattuna erilaisia variaatioita, joilla voi kehittää pikajuoksijan voimantuotto-ominaisuuksia. Näissä kahdessa artikkelissa on käyty läpi suhteellisen kattavasti pikajuoksijan voimaharjoittelu teoria ja käytäntö. Koko paketin vetäminen kasaan, ohjelmointi, progression rakentaminen ja periodisaatio jää valmentajan harteille, jos siihen kaipaa apua niin ota yhteyttä. Tsempit reeneihin!
Lähteet:
Askling, C., Karlsson, J., & Thorstensson, A. (2003, August). Hamstring injury occurrence in elite soccer players after preseason strength training with eccentric overload. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/12859607
Baker, D., and Nance, S. The Relation Between Running Speed and Measures of Strength and Power in Professional Rugby League Players. J. Strength Cond. Res.13(3): 230-235, 1999.
Behrens, M., Mau-Moeller, A., Mueller, K., Heise, S., Gube, M., Beuster, N., … Bruhn, S. (2015, February 4). Plyometric training improves voluntary activation and strength during isometric, concentric and eccentric contractions. Retrieved from https://www.sciencedirect.com/science/article/pii/S1440244015000377
Bosch, F., and Klomp, R. Running: Biomechanics and Exercise Physiology Applied in Practice. Philadelphia, PA: Elsevier, 2005.
Čoh, M., & Zvan, M., Veličkovska, L., Zivkovic, V. & Gontarev, S. (2016). BIODYNAMICALFACTORSOFRUNNINGSPEEDDEVELOPMENT. 5. 17-22.
Colyer, S. L., Stokes, K. A., Bilzon, J. L. J., Holdcroft, D., & Salo, A. I. T. (2018, April 1). Training-Related Changes in Force-Power Profiles: Implications for the Skeleton Start. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28872389
Cross MR, Lahti J, Brown SR, Chedati M, Jimenez-Reyes P, Samozino P, et al. Training at maximal power in resisted sprinting: optimal load determination methodology and pilot results in team sport athletes. PLoS One. 2018;13(4):e0195477.
Cronin, J., Ogden, T., Lawton, T., and Brughelli, M. Does Increasing Maximum Strength Improve Sprint Running Performance. 29(3): 86-95, 2007.
Cunha, L., Alves, F., & Veloso, A. (2002). The touch-down and takeoff angles in different phases of 100 m sprint running. Presentation at the International Symposium on Biomechanics in Sport, Caceres-Extremadura, Spain.
Erskine, R. M., Jones, D. A., Maffulli, N., Williams, A. G., Stewart, C. E., & Degens, H. (2011, February). What causes in vivo muscle specific tension to increase following resistance training? Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/20889606
Haugen, T., Seiler, S., Sandbakk, Ø. et al. The Training and Development of Elite Sprint Performance: an Integration of Scientific and Best Practice Literature. Sports Med - Open5, 44 (2019). https://doi.org/10.1186/s40798-019-0221-0
Hucteau, E., Jubeau, M., Cornu, C. et al. Is there an intermuscular relationship in voluntary activation capacities and contractile kinetics?. Eur J Appl Physiol120, 513–526 (2020). https://doi.org/10.1007/s00421-019-04299-z
Janusevicius, D., Snieckus, A., Skurvydas, A., Silinskas, V., Trinkunas, E., Cadefau, J. A., & Kamandulis, S. (2017, June 1). Effects of High Velocity Elastic Band versus Heavy Resistance Training on Hamstring Strength, Activation, and Sprint Running Performance. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5465986/
Kanehisa H, Funato K, Kuno S, et al. Growth trend of the quadriceps femoris muscle in junior Olympic weight lifters: an 18-month follow-up survey. Eur J Appl Physiol 2003; 89: 238-42.
Kumagai K, Abe T, Brechue WF, et al. Sprint performance is related to muscle fascicle length in male 100-m sprinters. J Appl Physiol 2000; 88: 811-6.
Miller R, Balshaw TG, Massey GJ, Maeo S, Lanza MB, Johnston M, Allen SJ, Folland JP. The Muscle Morphology of Elite Sprint Running. Med Sci Sports Exerc. 2020 Oct 1. doi: 10.1249/MSS.0000000000002522. Epub ahead of print. PMID: 33009196.
Morin, J.-B., Petrakos, G., Jiménez-Reyes, P., Brown, S. R., Samozino, P., & Cross, M. R. (2017, July). Very-Heavy Sled Training for Improving Horizontal-Force Output in Soccer Players. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/27834560
Morin JB, Petrakos G, Jiménez-Reyes P, Brown SR, Samozino P, Cross MR. Very-heavy sled training for improving horizontal-force output in soccer players. Int J Sports Physiol Perform. 2017;12(6):840–4.
Nagahara R, Zushi K. Development of maximal speed sprinting performance with changes in vertical, leg and joint stiffness. J Sports Med Phys Fitness. 2017 Dec;57(12):1572-1578. doi: 10.23736/S0022-4707.16.06622-6. Epub 2016 Jul 13. PMID: 27406013.
Nagano, A., & Komura, T. (2003, November). Longer moment arm results in smaller joint moment development, power and work outputs in fast motions. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/14522209
Petrakos G, Morin JB, Egan B. Resisted sled sprint training to improve sprint performance: a systematic review. Sports Med. 2016;46(3):381–400.
Poliquin, C., Patterson, Paul. Terminology: Classification of Strength Qualities. Strength Conditioning J. 11(6):48-52, 1989.
Reich, T. E., Lindstedt, S. L., LaStayo, P. C., & Pierotti, D. J. (2000, June). Is the spring quality of muscle plastic? Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/10848536
Ross, S. A., & Wakeling, J. M. (2016, June). Muscle shortening velocity depends on tissue inertia and level of activation during submaximal contractions. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4938035/
Schache, A. G., Blanch, P. D., Dorn, T. W., Brown, N. A. T., Rosemond, D., & Pandy, M. G. (2011, July). Effect of running speed on lower limb joint kinetics. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/21131859
Widrick, J. J., Stelzer, J. E., Shoepe, T. C., & Garner, D. P. (2002, August). Functional properties of human muscle fibers after short-term resistance exercise training. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/12121854
Wilson, G., Newton, R., Murphy, A. & Humphries, B. (1993) The optimal training load for the development of dynamic athletic performance. Med Sci Sports Exerc. Nov;25(11):1279-86. PMID: 8289617.
Perinteinen hidas voimaharjoittelu ei aiheuta pikajuoksijaa kehittäviä adaptaatioita
Lonkan ojentajille ja koukistajille voimaharjoittelua nopealla voimantuotolla
Reiden ojentajille ja koukistajille harjoittelussa tulee keskittyä taas korkeampaan voimantuottokykyyn, koska juoksussa polven alueen lihakset mahdollistavat korkean juoksunopeuden vastaanottamalla suuria voimia eksentrisesti.
Nilkka vastaa suurelta osin kontaktivaiheen voimantuotosta. Nilkan vahvistaminen pikajuoksun omaisilla harjoitteilla on tärkeää.
Käytännössä: samassa harjoituksessa ensiksi korkean nopeuden liikkeitä lonkan alueella ja tämän jälkeen eksentristä harjoittelua polven alueen lihaksille.
”Voimaharjoittelu tekee hitaaksi” - on suhteellisen yleisesti kuultu lausahdus. Nykyään tämä myytti on jo onneksi purettu ja voimaharjoittelua käytetään enemmän myös nopeuden kehittämisessä, mutta mikä on oikea määrä ja tapa käyttää voimaharjoittelua, jos tavoitteena on maksimaalisen nopeuden kehittäminen, eikä pelkästään alkukiihdyttämisen parantaminen?
Pikajuoksijan, eli 100–200 metrin juoksijan, voimaharjoittelu ei ole niin yksinkertaista kuin sen voisi kuvitella olevan. Voimaharjoittelu kehittää lähes suoraan räjähtävämmän startin ja paremman kiihdytyksen, mutta entä voiko voimaharjoittelulla vaikuttaa maksimijuoksuvaiheeseen ja maksimijuoksunopeuteen?
Pikajuoksu ja voiman eri lajit
Juoksu voidaan jakaa karkeasti kiihdyttämiseen ja maksimijuoksuvaiheeseen. Monessa tutkimuksessa on löydetty voimaharjoittelun ja kiihdyttämisen yhteys, mutta voimaharjoittelun yhteys maksimijuoksuvaiheeseen on aiheuttanut ristiriitaisia tutkimustuloksia (esim. Wilson et al., 1993).
Pikajuoksija tarvitsee pääasiassa nopeusvoimaa. Nopeusvoiman voi määritellä hermolihasjärjestelmän kyvyksi tuottaa mahdollisimman paljon voimaa lyhyimmässä mahdollisimmassa ajassa (Poliquin & Patterson, 1989). Pikajuoksussa nopeusvoimasuorituskykyä voi parantaa kehittämällä voimantuottonopeutta, maksimivoimatasoja tai kumpaakin samaan aikaan. Pikajuoksija tarvitsee siis tietyn verran maksimivoimaakin. Maksimivoimaa kehittääkseen voi olla järkevä jossain määrin kehittää myös perusvoimaa ja kenties lihasten kokoa. Lisäksi tarvitaan nopeusvoiman kohdalla erilaisia ominaisuuksia, kuten:
Lähtövoimaa
Räjähtävää voimaa
Reaktiivista voimaa
Lähtövoima tarkoittaa voiman tuottamista staattisesta asennosta ja kehonpainomme inertian ylittämistä. Tätä tarvitaan erityisesti ensimmäisessä askeleessa. Onkin hyvin loogista, että voimaharjoittelu kehittää juuri ensimmäisiä askelia huomattavasti.
Räjähtävä voima taas on kykyä jatkuvasti kasvattaa voimaa aikayksikkö kohden, kun liike on jo aloitettu. Reaktiivinen voima tarkoittaa taas kehon kykyä vaihtaa nopeasti eksentrisestä supistuksesta konsentriseen supistukseen. Tässä kyvyssä korostuu erityisesti hermolihasjärjestelmän kyky maksimoida venymis-lyhenemis-syklus. Reaktiivinen voima kehittyy myös, kun lihaksen jäykkyys lisääntyy ja elastiset ominaisuudet kehittyvät. (Bosch & Klomp, 2005; Siff, 2003; Poliquin & Patterson, 1989.) Harjoituksellisesti voimme vielä jakaa edellä mainitut voimat eksentrisiin, konsentrisiin ja isometrisiin komponentteihin.
Kuva 1: Juoksussa rasittuvat erityisesti pohkeet, takareidet, pakarat ja selän ojentajat. Lisäksi vatsalihakset ja olkapäät tekevät kovasti töitä juoksun rytmityksen ja asennon hallinnan kanssa. Juoksun kontaktivaiheessa erityisesti pohjelihas tekee kovasti töitä (Čoh et al., 2016)
Lähdössä tarvitaan konsentrista voimantuottoa – maksimijuoksuvaiheessa reaktiivista voimaa
Maksimaalisella voimalla ja 10 m sekä 30 m juoksuajoilla ei ole merkitsevää yhteyttä, mutta kun yhtälöön lisätään juoksijan kehonpaino, niin löydetään tilastollisesti merkitsevä yhteys (Baker et al., 1999; Cronin et al., 2007). Bakerin ja kumppaneiden tutkimuksessa myös huomattiin, että 10 m tulos oli yhteydessä suhteelliseen voimantuottoon ja konsentriseen voimantuotto kykyihin. Lähdössä konsentrisen voiman kehittämisestä on hyötyä, koska merkitsevää venymislyhenemis-syklusta ei tapahdu. Lisäksi lähdössä kontaktiaika on suurempi kuin juoksun muissa vaiheissa ja tämä mahdollistaa suuremman voiman hyödyntämisen askelkontaktin aikana (taulukko 1).
TAULUKKO 1: Keskiarvolliset kinemaattiset muuttujat 100 metrin juoksussa hyvin harjoitelleilla pikajuoksijoilla (Cunha et al., 2002).
Juoksun vaihe
Nopeus (m/s)
Askelpituus (m)
Askelfrekvenssi (Hz)
Kontaktiaika (s)
Lentoaika (s)
Kiihdytys
9,80
4,16
2,36
0,10
0,12
Maksimijuoksuvaihe
10,46
4,48
2,34
0,08
0,13
Ylläpito
9,85
4,36
2,26
0,10
0,13
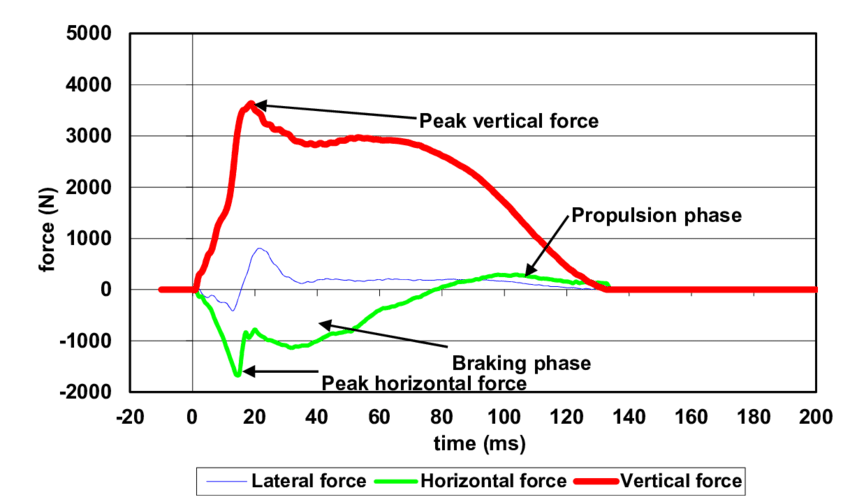
Maksimaalinen juoksu taas yhdistetään pystympään juoksuasentoon, jonka tarkoituksena on maksimoida askelpituus, askelfrekvenssi ja minimoida kontaktiaika. Tämä pystympi juoksuasento johtaa suurempiin verikaalisiin ja jarruttaviin voimiin verrattuna kiihdytysvaiheeseen (kuva 2). Nämä voimat tuottavat suuria venytysvoimia lihaksiin ja näin luovat isomman tarpeen venymislyhenemis kyvyille lihaksissa. Jotta näitä voimia voidaan hyödyntää maksimaalisesti, tulee kehittää elastista/reaktiivista voimaa. Lihaksen kyky kontrolloida suuria voimia eksentrisissä tai isometrisissä supistuksissa on äärimmäisen tärkeä harjoitettava ominaisuus. Ilman sitä reaktiivinen voima ei voi kehittyä.
Kuva 2: Kontaktivaiheessa vertikaalivoimat korostuvat (Čoh et al., 2016).
Voimaharjoittelu ensiksi ja sitten nopeus - ajattelu romukoppaan
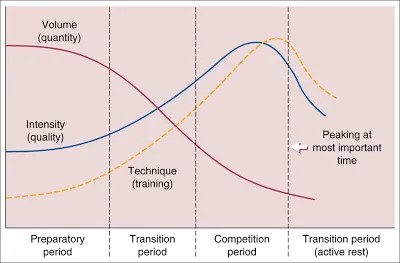
Pikajuoksijat tekevät yleensä voimaharjoittelussaan kaikille lihaksille samanlaista voimaa. Tämä ei tuota optimaalisinta lopputulosta ja saattaa jopa hidastaa kokeneita urheilijoita. Pikajuoksijan voimaharjoittelun periodisaatio perustuu väärille olettamuksille. Perinteisesti ensiksi voimaharjoittelulla nostetaan maksimivoimareserviä. Tämän jälkeen tehdään voimaharjoittelua lyhyillä voimantuottoajoilla. Tavoitteena saada kasvanut maksimivoimareservi käyttöön jälkimmäisellä jaksolla (kuva 3). Tämä ei ole tehokas tapa kokeneelle urheilijalle.
Kuva 3: Perinteisen periodisaation mukaan on ensiksi tarkoitus opetella tuottamaan paljon voimaa. Tämän jälkeen yritetään tuottaa samaa voimaa korkealla nopeudella, eli tehdään enemmän spesifiä voimaharjoittelua ja yritetään saada voima siirtymään lajiin. Tällöin perinteisen voimaharjoittelun osuus pienenee.
Ajatus siitä, että voimaharjoittelu tuottaa pohjan, jotta nopeutta voidaan kehittää, on vääristynyt. Välttämättä tällaista yleistä voimaharjoittelujaksoa ei tarvita ollenkaan. Tämä johtuu siitä, että kova voimaharjoittelu tuottaa kolmenlaista tärkeää adaptaatiota:
Motorisen yksikön rekrytointi paranee
Lateraalisen voiman siirtyminen paranee
Lihas kasvaa, eli tapahtuu hypertrofiaa
Nämä adaptaatiot eivät ole tärkeitä pikajuoksun kannalta ja tarvittavat adaptaatiot voidaan saavuttaa muullakin tavalla kuin perinteisellä hitaalla voimaharjoittelulla.
Ensimmäisen adaptaation, eli motorisen yksikön rekrytoimista voidaan kehittää myös korkeilla voimantuottonopeuksilla (Behrens et al., 2015). Sen kehittämiseen ei tarvita välttämättä ollenkaan perinteistä voimaharjoittelua. Toisen adaptaation, eli parantuneen lateraalisen voimantuoton siirtyminen ei paranna nopeaa voimantuottoa, joten sekään ei ole tärkeä pikajuoksun kannalta (Erskine et al., 2011).
Kolmannella adaptaatiolla, eli hypertrofialla on merkityksensä nopeaan voimantuottoon, mutta siihen liittyy muutama pohdittava asia. Ensinnäkin hypertrofinen harjoittelu johtaa yleensä nopeimman IIX lihastyypin muuttumiseksi IIA lihassoluksi. IIA on hitaampi lihassolu, joten tämä ei ole haluttavaa pikajuoksijalla. (Widrick et al., 2002.)
Toiseksi hypertrofinen harjoittelu johtaa lihaksenkasvuun ja näin sisäisen vipuvarren kasvuun, joka aiheuttaa sen, että lihassolujen pitää supistua vielä nopeammin, jotta saavutetaan sama vipuvarren kulmanopeus (Nagano & Komura, 2003). Tämä johtuu siitä, että lihaksen sisäisen vipuvarren kasvaessa pitää lihassolujen supistua pidemmälle saavuttaakseen saman nivelen liikelaajuuden ja ottaakseen kiinni tämän pidemmän matkan tulee lihassolujen supistua nopeammin saavuttaakseen saman nivelkulmanopeuden.
Kolmanneksi kudoksen inertia kasvaa, kun lihasmassa lisääntyy, mikä johtaa tietenkin hidastuneeseen nopeuteen (Ross & Wakelinf, 2016).
Viimeiseksi kova voimaharjoittelu johtaa lisääntyneeseen antagonistien (vastavaikuttajalihasten) koaktivaation nivelissä nopeissa liikkeissä. Tämä voi hidastaa juoksunopeutta. (Janusevicius et al., 2017.) Tämä johtuu luultavasti siitä, että keho yrittää suojella isompaa lihasta vaurioilta aktivoimalla vastavaikuttajalihasta, mikä hidastaa päävaikuttajalihaksen supistumisnopeutta. Kovan voimaharjoittelun on näytetty vähentävän maksimaalista nopeutta (Colyer et al., 2018) mm. edellä mainittujen seikkojen takia, mutta myös voimaharjoittelun aiheuttaman väsymyksen takia.
Joskus hypertrofialla on paikkansa isompaa kokonaisuutta rakentaessa, mutta eivätkö nopeat liikkeet aiheuttaisi hypertrofisia adaptaatioita, jos ne olisivat elintärkeitä nopealla liikkeelle? Näin ei ikävä kyllä ole ja sen takia hypertrofista harjoittelua pikajuoksijalle tulee punnita tarkkaan.
Vielä lisäyksenä, että etu- ja takareisille on pystytty saamaan lisää eksentristä voimaa ilman kovaa perinteistä voimaharjoittelua pelkästään tekemällä eksentristä voimaharjoittelua (Reich et al., 2000). Nämä adaptaatiot ovat luultavimmin johtuneet titinin ja kollageenin adaptoitumisesta lihaksen sisällä.
Aloittelijoille toimii perinteinen voimaharjoittelu – kokeneille ei
Perinteinen voimaharjoittelu parantaa voluntaarista aktivointikykyä, minkä kehittäminen parantaa voimantuottonopeutta ja maksimivoimaa. Yleensä aloittelijoilla ei ole kykyä aktivoida kaikkia lihassoluja, joita he kontrolloivat. Voimaharjoittelun jälkeen tämä kyky paranee ja lisää motorisia yksiköitä tulee saataville. Tämä parantaa maksimivoimaa ja voimantuottokykyä. Mutta kokeneilla harjoittelijoilla tätä adaptaatioita ei enää saavuteta ja voluntaarisen aktivointikyvyn parantaminen on erittäin hankalaa. (Hucteau et al., 2020.)
Voimaharjoittelu parantaa reisien eksentristä voimaa ja lonkan ojentajien voimaa myös korkeilla nopeuksilla, vaikkakin ei yhtä paljon kuin pelkkä nopea voimantuottoharjoittelu. Voimaharjoittelun teho pienenee, kun urheilija saavuttaa riittävän voimatason tai maksimaalisen konsentrisen supistumisnopeuden. Lisäksi voimaharjoittelu saattaa kääntyä itseään vastaan, jos kehonpainon on lisäännyttävä, jotta lihasmassa lisääntyisi. Näitä ongelmia ei kohdata heti aluksi ja sen takia voimaharjoittelu on aluksi tehokasta. Kokeneempien urheilijoiden tulee miettiä muita vaihtoehtoja.
Minkälaista voimaharjoittelun pitäisi sitten olla pikajuoksijalle?
Pikajuoksijan kannattaa keskittyä elastisen voiman kehittämiseen. Elastinen voima tarkoittaa kudosten kykyä imeä, varastoida ja vapauttaa energiaa. Mitä enemmän energiaa nämä kudokset vapauttavat, sitä nopeammin ja tehokkaammin juoksija kiitää radalla. Elastinen energia tuotetaan jänteellä. Esimerkiksi pohjelihas ei veny juoksussa eksentrisessä vaiheessa, vaan lihas pysyy saman pituisena (tekee isometristä työtä) ja jänne venyy, mutta koko lihasjännekompleksi kasvaa. Jänteet ovat kuin jousia ja venytettäessä ne kimmahtavat nopeasti takaisin lepopituuteensa. Jänteet tuottavat siis paljon ylimääräistä nopeaa voimaa. Jänteet eivät tarvitse happea ja eivätkä väsy. Jänteiden ja elastisten ominaisuuksien kehittäminen olisi tärkeä olla mukana kaikessa tekemisessä.
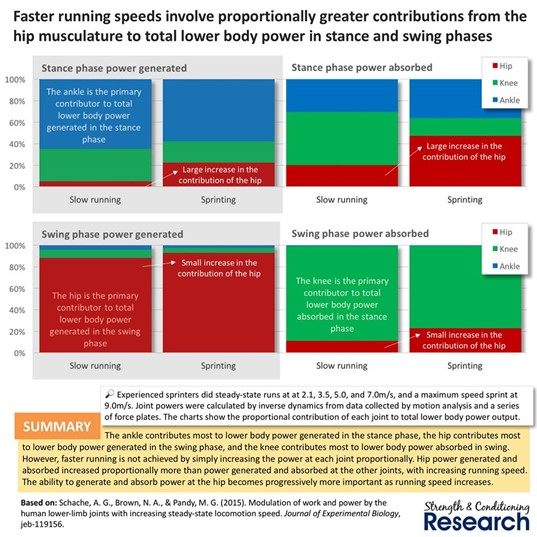
Voimaharjoittelussa lihaksia tulisi kehittää pikajuoksun tarpeiden mukaan. Tukivaiheessa, eli toisen jalan ollessa maassa, nilkka vastaa suurimmalta osalta voimantuotosta. Nilkan ja nilkkaa liikuttavien lihasten voimaharjoittelu on erittäin tärkeää. Jalan heilahdusvaiheessa eniten voimaa tuottaa taas lantion alueen lihakset. Eniten voimaa imevät polven alueen lihakset: taka- ja etureisi. Juoksunopeuden lisääntyessä kyky tuottaa ja ottaa vastaan voimia erityisesti lantion alueella tulee progressiivisesti tärkeämmäksi (Kuva 4).
Pakaroihin, lähentäjiin, takareisiin ja lonkan koukistajiin tarvitaan nopeaa voimantuottoa erityisesti heilahdusvaiheessa! Etureidet ja takareidet taas imevät eniten voimaa ja toimivat tärkeinä tekijöinä tukivaiheessa. Tärkeimmät lihakset tukivaiheessa ovat soleus ja gastrocnemius sekä erityisesti liikesuuntana nilkan plantaarifleksio. (Schache et al., 2011.) Huomiona kuitenkin, ettei voimantuotto tukivaiheessa ole yhteydessä juoksunopeuden kanssa. Se toimii juoksunopeuden mahdollistavana tekijänä.
Joten tarvitaan nopeaa voimantuottoa lonkan ojentajiin ja koukistajiin sekä eksentristä voimaa polvien ojentajien ja koukistajien, jotta ne voivat imeä voimia!
Heilautusvaiheessa nopea supistumisnopeus, korkea nopeus --> nopean voimantuoton harjoittelua lonkan alueelle!
Kun taas etureidet heilautusvaiheessa imevät voimaa niiden lihaspituus kasvaa! Tällöin nopeudella ei ole väliä vaan voimantuottokyvyllä. (Askling et al., 2003.)
Voimaharjoittelua tulisi tehdä siis lonkan ojentajille ja koukistajille korkeilla nopeuksilla. Reiden ojentajille ja koukistajille taas korkeampaa voimantuottoa, kun niiden pituus kasvaa. Tämä johtuu siitä, että niiden tärkein tehtävä juoksun aikana on ottaa vastaan isoja voimia. Käytännössä tämä voisi tarkoittaa sitä, että samassa harjoituksessa tehtäisiin ensiksi korkean nopeuden liikkeitä lonkan alueella ja tämän jälkeen eksentristä harjoittelua polven alueen lihaksille.
Korkean nopeuden voimaharjoittelu parantaa korkean nopeuden voimaa paljon enemmän kuin raskas perinteinen voimaharjoittelu ja eksentrinen harjoittelu parantaa eksentristä voimaa paljon enemmän kuin perinteinen voimaharjoittelu. Tämän takia spesifimmän harjoittelun rakentaminen on paljon tehokkaampaa kuin perinteisen voimaharjoittelu tekeminen. Korkean voiman ja korkean nopeuden periodisointi peräkkäin ei ole järkevää pikajuoksussa, koska eri lihakset tarvitsevat erilaista voimantuottokykyä. Niitä voi ja kannattaa kehittää kokoajan! Lisäksi hyötynä tässä on se, ettei tarvitse pelätä jonkun ominaisuuden laskevan, kuten perinteisessä periodisaatiossa. Esimerkiksi perinteisessä periodisaatiossa reisien voimantuotto tippuu, kun siirrytään harjoittamaan nopeaa voimantuottoa.
Kuva 4: Nilkka tuottaa voimaa kontaktivaiheessa kun taas heilahdusvaiheessa voimantuotosta vastaa lantio. Voimia imee polven alueen lihakset kontaktivaiheessa, mutta myös lantion alueen lihakset vauhdin kasvaessa.
Lähteet:
Askling, C., Karlsson, J., & Thorstensson, A. (2003, August). Hamstring injury occurrence in elite soccer players after preseason strength training with eccentric overload. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/12859607
Baker, D., and Nance, S. The Relation Between Running Speed and Measures of Strength and Power in Professional Rugby League Players. J. Strength Cond. Res.13(3): 230-235, 1999.
Behrens, M., Mau-Moeller, A., Mueller, K., Heise, S., Gube, M., Beuster, N., … Bruhn, S. (2015, February 4). Plyometric training improves voluntary activation and strength during isometric, concentric and eccentric contractions. Retrieved from https://www.sciencedirect.com/science/article/pii/S1440244015000377
Bosch, F., and Klomp, R. Running: Biomechanics and Exercise Physiology Applied in Practice. Philadelphia, PA: Elsevier, 2005.
Čoh, M., & Zvan, M., Veličkovska, L., Zivkovic, V. & Gontarev, S. (2016). BIODYNAMICALFACTORSOFRUNNINGSPEEDDEVELOPMENT. 5. 17-22.
Colyer, S. L., Stokes, K. A., Bilzon, J. L. J., Holdcroft, D., & Salo, A. I. T. (2018, April 1). Training-Related Changes in Force-Power Profiles: Implications for the Skeleton Start. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28872389
Cronin, J., Ogden, T., Lawton, T., and Brughelli, M. Does Increasing Maximum Strength Improve Sprint Running Performance. 29(3): 86-95, 2007.
Cunha, L., Alves, F., & Veloso, A. (2002). The touch-down and takeoff angles in different phases of 100 m sprint running. Presentation at the International Symposium on Biomechanics in Sport, Caceres-Extremadura, Spain.
Erskine, R. M., Jones, D. A., Maffulli, N., Williams, A. G., Stewart, C. E., & Degens, H. (2011, February). What causes in vivo muscle specific tension to increase following resistance training? Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/20889606
Hucteau, E., Jubeau, M., Cornu, C. et al. Is there an intermuscular relationship in voluntary activation capacities and contractile kinetics?. Eur J Appl Physiol 120,513–526 (2020). https://doi.org/10.1007/s00421-019-04299-z
Janusevicius, D., Snieckus, A., Skurvydas, A., Silinskas, V., Trinkunas, E., Cadefau, J. A., & Kamandulis, S. (2017, June 1). Effects of High Velocity Elastic Band versus Heavy Resistance Training on Hamstring Strength, Activation, and Sprint Running Performance. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5465986/
Kanehisa H, Funato K, Kuno S, et al. Growth trend of the quadriceps femoris muscle in junior Olympic weight lifters: an 18-month follow-up survey. Eur J Appl Physiol 2003; 89: 238-42.
Kumagai K, Abe T, Brechue WF, et al. Sprint performance is related to muscle fascicle length in male 100-m sprinters. J Appl Physiol 2000; 88: 811-6.
Morin, J.-B., Petrakos, G., Jiménez-Reyes, P., Brown, S. R., Samozino, P., & Cross, M. R. (2017, July). Very-Heavy Sled Training for Improving Horizontal-Force Output in Soccer Players. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/27834560
Nagahara R, Zushi K. Development of maximal speed sprinting performance with changes in vertical, leg and joint stiffness. J Sports Med Phys Fitness. 2017 Dec;57(12):1572-1578. doi: 10.23736/S0022-4707.16.06622-6. Epub 2016 Jul 13. PMID: 27406013.
Nagano, A., & Komura, T. (2003, November). Longer moment arm results in smaller joint moment development, power and work outputs in fast motions. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/14522209
Poliquin, C., Patterson, Paul. Terminology: Classification of Strength Qualities. Strength Conditioning J. 11(6):48-52, 1989.
Reich, T. E., Lindstedt, S. L., LaStayo, P. C., & Pierotti, D. J. (2000, June). Is the spring quality of muscle plastic? Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/10848536
Ross, S. A., & Wakeling, J. M. (2016, June). Muscle shortening velocity depends on tissue inertia and level of activation during submaximal contractions. Retrieved from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4938035/
Schache, A. G., Blanch, P. D., Dorn, T. W., Brown, N. A. T., Rosemond, D., & Pandy, M. G. (2011, July). Effect of running speed on lower limb joint kinetics. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/21131859
Widrick, J. J., Stelzer, J. E., Shoepe, T. C., & Garner, D. P. (2002, August). Functional properties of human muscle fibers after short-term resistance exercise training. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/12121854
Wilson, G., Newton, R., Murphy, A. & Humphries, B. (1993) The optimal training load for the development of dynamic athletic performance. Med Sci Sports Exerc. Nov;25(11):1279-86. PMID: 8289617.
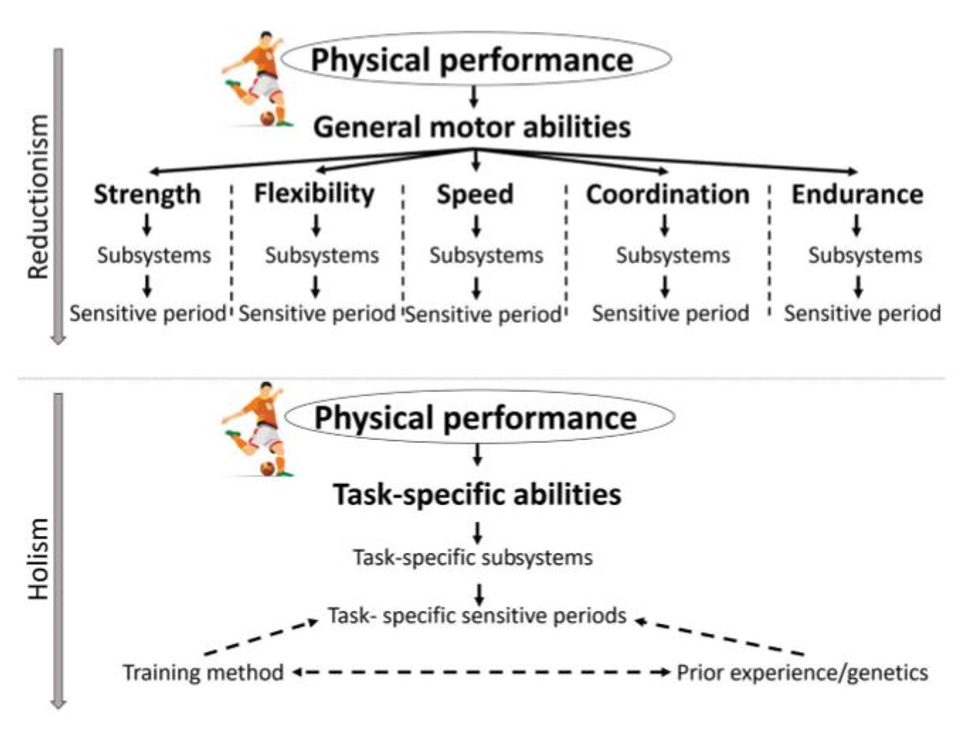
Suurin osa valmennuskirjallisuudesta viittaa yleisiin motorisiin ominaisuuksiin. Näillä tarkoitetaan yleensä voimaa, nopeutta, kestävyyttä, liikkuvuutta ja taitavuutta. Näiden väitetään olevan yläkäsitteitä, joiden siirtovaikutus on erittäin hyvä käsitteen alla. Esimerkiksi pelkästään nopeus voidaan jakaa lukuisiin eri alalajeihin, joissa toisen kehittäminen ei kehitä toista. Yleistaidot eivät ole erikseen harjoitettavissa, vaikka niin väitetään. Olisiko yleisten motoristen ominaisuuksien sijaan tarkastelussa siirryttävä tarkemmin yksittäisiin motorisiin taitoihin?
Lähekkäin olevat taidot eivät korreloi keskenään kovinkaan vahvasti. Näyttäisi siltä, että on monia erilaisia motorisia malleja ja ei ole yhtä yleistaitoja, jonka alle nämä voisivat rakentua. Esimerkiksi Drowatzky & Zuccato (1967) testasivat kuutta eri tasapainotestiä ja miten koehenkilöt pärjäävät näissä testeissä. Jos tasapaino yleistaito olisi yksi malli niin, silloin yhdessä testissä pärjääminen olisi yhteydessä pärjäämiseen hyvin kaikissa testeissä. Kuitenkin tuloksissa korkein korrelaatio oli vaatimaton r=0.31 tarkoittaen vain noin 9,6 % yhteyttä testien välillä! Valmentamisessa keskittymisen tulisikin olla enemmän yksittäisissä motorisissa taidoissa kuin yleistaidoissa. Miten näitä tulisi sitten harjoittaa?
Yleensä harjoittelussa valmentaja antaa kognitiivisella tasolla ohjeita urheilijalle/urheilijoille. Urheilija yrittää kuunnella ja sisäistää asian ja sen jälkeen tietoisesti korjata virheellisen suorituksen. Taitojen harjoitteleminen tietoisella tasolla on aivan liian hidasta. Pelissä ja suorituksessa refleksit korjaavat eniten liikettä, joten harjoittelunkin tulisi olla samanlaista!
Nopeat liikkeet suunnitellaan etukäteen ja sitten ne vain toteutetaan. Keholla ei ole aikaa prosessoida kaikkea palautetta virheistä ja korjata niitä suorituksen aikana, vaan liikkeet toteutetaan ennalta rakennetun mallin mukaan. Liikkeen aikana on liikaa liikkuvia lihaksia ja niveliä, joita pitää kontrolloida. Rajoittunut tietoinen tasomme ei millään pysty kontrolloimaan näitä kaikkia liikkeen aikana. Esimerkiksi uimahyppääjä ei pysty suorituksen aikana kontrolloimaan suuresti liikettään, vaan palaute liikkeestä tulee vasta liikkeen jälkeen. Esimerkiksi keihäänheittäjä voi pohtia heiton jälkeen miltäs se nyt tuntui ja mitäs siinä oikein tapahtui. Harjoittelulla voidaan rakentaa uusia motorisia ohjelmia tai parantaa vanhoja. Motorinen ohjelma sisältää aluksi mitä lihaksia käytetään, missä järjestyksessä ja kuinka pitkään.
Harjoittelulla saatetaan saavuttaa tila, missä joitakin refleksejäkin pystytään hallitsemaan, jotta suoritus olisi mahdollisimman hyvä. Aloittelevalla harjoittelijalla muodostuu paljon yksittäisiä erillisiä motorisia ohjelmia, jotka harjoittelun myötä sulautuvat yhdeksi kokonaiseksi motoriseksi ohjelmaksi. Siksi huipuilla taito näyttää niin helpolta ja sujuvalta. Kun aloittelevalla se voi olla vähän kulmikas ja töksähtelevä. Tämä johtuu siitä, että aina pitää aktivoida uusi motorinen ohjelma ja se on kömpelöä.
Tässä näkyy hyvin, miten tavoite on osua palloon ja se ei onnistu, jos on paljon liikkuvia osia. Sen takia motorinen ohjelma sulkee esimerkiksi selän kierron ja lonkkanivelen liikkeen videossa, jotta tavoite onnistuu. Onnistuakseen tavoitteessa, eli palloon osumisesta, tulee kehon vähentää nivelten vapaata liikkumista. Tätä voidaan kutsua liikkeen jäädyttämiseksi.
Ei se aina entisiltä huippu-urheilijoitakaan suju. Tämä kertoo eri motorisen liikemallin olevan käytössä golfissa kuin koripallossa, eivätkä nämä mene yhden ”yleistaidon” alle.
Taidon harjoittelu auttaa motorista ohjelmaa kehittymään ja lisää vapausastetta sekä liikkeeseen tulee enemmän liikkuvia osia, jotka lopulta näkyvät taidokkaampana lyöntinä. Tavoite pysyy koko ajan samana. Osua palloon ja saada se johonkin haluttuun kohtaan tippumaan. Bernstein (1967) kutsuu tätä ensimmäistä oppimisen vaihetta vapausasteen ongelmaksi. Kaikki kehon eri lihakset ja nivelet ovat vapaita liikkuman useaan eri suuntaan, Oppija ei vielä osaa kontrolloida kaikkea tätä vapautta ja siten keho vähentää ei-tärkeimmät kehon osat pois aloittelevalta taidon oppijalta, jotta tavoitteessa onnistutaan.
Myöhemmin oppija voi ruveta ottamaan mukaan enemmän vapausastetta, jotka olivat aluksi “jäätyneitä”.
Tämä mahdollistaa nopeamman ja suuremman voimantuoton varsinkin nopeissa liikkeissä (kuva 2). Kaikkein kokeneimmat urheilijat oppivat Bernsteinin mukaan hyödyntämään kehon passiivisia elementtejä, kuten momentumia, elastisuutta, painovoimaa ym. Tämä johtaa tehokkaampaan liikkeen suorittamiseen ja pienempään energian tuhlaukseen. Esimerkiksi lämääminen jääkiekossa vaatii mailan elastisuuden hyödyntämistä ja esimerkiksi aloittelevat jääkiekkopelaajat eivät opi ensimmäiseksi lämäämistä.
KUVA 2: Hyvä esimerkki aloittelijan ja kokeneen taidon eroista. Vapausasteen ero näkyy ylävartalon kierrossa. Oikealla puolella kohta pääse lantio työntymään eteen ja päästään hyödyntämään venytysrefleksin tuomaa lisävoimaa ja ylävartalon kiertovoiomaa, kun taas vasemmalla voidaan voimaa tuottaa enää työntökädellä. Ylävartaloon ei ole muodostunut kiertoa, koska se vaatisi liikaa vapausastetta yläselän nikamilta, joka hankaloittaisi tavoitetta, eli kuulan työntämistä pois ringistä.
Rytmi on motoristen ohjelmien pohja
Nopeat liikkeet ovat jokainen erilaisia, vaikka motorinen ohjelma taustalla olisi sama. Schöllhorn seurasi yhden koko vuoden ajan toistaisiko kaksi huippukiekonheittäjä heittoliikkeensä täsmälleen samanlaisena (Savelsbergh et al, 2010). Vuoden aikana urheilijat eivät toistaneet kertaakaan samaa heittoa kahdesti. Liike ei ole koskaan täydellisesti samanlainen, vaikka usein tavoitellaankin sitä yhtä tiettyä täydellistä tekniikka. Motorinen ohjelma sovelletaan aina eri ympäristöön ja tilanteisiin. Esimerkiksi sulkapallolyönti ei ikinä tule samaan kohtaan tai lyöjän asento ei ole ikinä täsmälleen sama. Motorista ohjelmaa joudutaan aina hiukan varioimaan. Taito varioidaan ympäristön tarpeisiin. Motorinen ohjelma ilmaistaan eri tavalla, mutta sen perusluonne ei muutu. Yleinen motorinen ohjelma on pitkäaikaisessa muistissa. Sieltä se kaivetaan esiin ja sitä pystytään varioimaan eri tilanteissa.
Kuitenkin rytmi tai relatiivinen ajoitus toimii pohjana motorisille ohjelmille ja on vakio, vaikka muuten ympäristö vaihtuisi paljonkin. Liikkeen nopeuskin voi muuttua samassa motorisessa ohjelmassa. Nopeus vaikuttaa muun muassa voimiin, voimantuoton nopeuteen, raajojen nopeuteen ja kuljettuun matkaan ym., mutta rytmi on sama, vaikka nopeus kasvaisikin. Rytmi toimii motorisen ohjelman pohjana ja kaikkea muuta on helppo varioida sen ympärille. Tarkemmin hitaammin tehty liike hidastetaan kokonaisuutena hitaammaksi yksiköissä, jotta liikkeen suhteellinen ajoitus pysyy samana. Esimerkiksi lyhyempi ja pidempi heitto. Joten kokonainen yli olan tapahtuva heittäminen voidaan säilöä pitkäaikaismuistiin yhtenä kokonaisuutena. (Schmidt & Lee, 2011.)
KUVA 3: Loistava kuva demonstroimaan heittotaidon variointia eri välineellä. Nilkan, polven, lantion, hartian ajoitus on tässä tärkeä kuin myös keskivartalon kääntymisen ajoitus.
Mitä kaikkea voi sitten vaihdella motorisen ohjelman sisällä?
- Lihakset. Niin uskomattomalta kuin se kuulostaa joissain määrin lihaksien vaihtaminen onnistuu. Raibert (1977) tutki asiaa kirjoittamalla saman lauseen ensiksi käsillä, sitten ranne immobilisoituna, vasemmalla kädellä, hampailla ja viimeiseksi varpailla. Kaikki kirjoitukset pystyttiin selvästi tunnistamaan saman henkilön kirjoittamaksi, vaikka eri lihakset työskentelivät! Sama motorinen ohjelma oli käytössä, vaikka eri lihakset työskentelivät.
Joten mahdollistatko sinä valmennuksessasi motorisen ohjelman optimaalisen kehittymisen? Etenetkö opetuksessa järkevästi liikkeen taitotaso huomioiden? Lisäksi huomioitko liikkeen rytmin nopeiden liikkeiden ytimenä ja varioit taitoa loputtomasti, mutta pitäen liikkeen ytimen samana?
Lähteet:
Bernstein, N.A. (1967) The Co-ordination and regulation of movements. Oxford: Pergamon Press.
Drowatzky, J. & Zuccato, F. (1967) Interrelationships between Selected Measures of Static and Dynamic Balance, Research Quarterly. American Association for Health, Physical Education and Recreation, 38:3, 509-510, DOI: 10.1080/10671188.1967.10613424
Hooren, B. & Croix, M. (2020). Sensitive Periods to Train General Motor Abilities in Children and Adolescents: Do They Exist? A Critical Appraisal. Strength and Conditioning Journal. 1. 10.1519/SSC.0000000000000545.
Keele, S. W., & Posner, M. I. (1968). Processing visual feedback in rapid movements. Journal of Experimental Psychology, 77, 155-158. doi:10.1037/h0025754
Kernodle, M. & Carlton, L. (1992). Information Feedback and the Learning of Multiple-Degree-of-Freedom Activities. Journal of motor behavior. 24. 187-96. 10.1080/00222895.1992.9941614.
Lee, T. D., & Magill, R. A. (1983). The locus of contextual interference in motor-skill acquisition. Journal of Experimental Psychology: Learning, Memory, and Cognition, 9(4), 730-746. http://dx.doi.org/10.1037/0278-7393.9.4.730
McCracken, H. & Stelmach, G. (1977). A Test of the Schema Theory of Discrete Motor Learning. Journal of Motor Behavior. 9. 193-201. 10.1080/00222895.1977.10735109.
Raibert, M.H. (1977) Motor control and learning by the state space model. Tech. Rep. No. AI-TR-439. Cambridge: MIT, Artificial Intelligence Laboratory.
Rothstein A. L., Arnold R. (1976). Bridging the gap: application of research on videotape feedback and bowling. Mot. Skills Theory Pract. 1, 35–64
Ryan, R. M. & Deci, E. L. (2000). Self-determination theory and the facilitation of intrinsic motivation, social development, and well-being. American Psychologist, 55(1), 68-78.
Shea, J. & Zimny, S. (1983). Context Effects in Memory and Learning Movement Information. Res Q Exerc Sport. 1991 Jun;62(2):187-95. DOI: 10.1016/S0166-4115(08)61998-6.
Schmidt, R.A., & Lee, T.D (2011) Motor control and learning: A behavioral emphasis (5th edition). Champaign, IL: Human Kinetics.
Schmidt, R.A., & Young, D.E. (1987) Transefer of movement control in motor learning. In S.M: Cormier & J.D. Hagman (editors), Transfer of learning (p. 45-80). Orlando, FL: Academic press.
Schmidt, R. A. (1975). A schema theory of discrete motor skill learning. Psychological Review, 82(4), 225-260. http://dx.doi.org/10.1037/h0076770
Swinnen, S., Schmidt, R., Nicholson, D. & C. Shapiro, D. (1990). Information Feedback for Skill Acquisition: Instantaneous Knowledge of Results Degrades Learning. Journal of Experimental Psychology: Learning, Memory, and Cognition. 16. 706-716. 10.1037//0278-7393.16.4.706.
Ungerleider, L.G. & Mishkin, M. (1982) Two cortical visual systems. In D.K.Ingle, M.A. Goodale, & R.J.W. Mansfield (editors), Analysis of visual behaviour, pp. 549 - 587. Cambridge, MA: MIT Press.
Winstein, C. J., & Schmidt, R. A. (1990). Reduced frequency of knowledge of results enhances motor skill learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16(4), 677-691.
Käy lukemassa vieraskirjoitus Lihastohtorin blogista miten lihasten kokoa ja voimaa voi ylläpitää sekä kehittää ilman pääsyä kuntosalille. Viime viikon harjoittelutauko kirjoituksen jatkoksi julkaistu käytännön osuus on nyt ulkona ja tarjoaa paljon käytännön vinkkejä kuvien ja videoiden muodossa.
Artikkelin yhteenveto:
Tällä hetkellä ei kannata keskittyä siihen mitä kaikkea ei pysty tekemään, vaan sen sijaan mitä pystyy tekemään.
Nyt tämä tilanne pakottaakin monet monipuolistamaan harjoittelua ja antaa mahdollisuuden oppia paljon uusia liikkeitä ja tapoja harjoitella.
Ilman kuntosalin laitteita ja välineitä voi harjoitella ja vähintäänkin ylläpitää saavutettuja voima- ja lihasmassatasoja.
Kehittävää tai vähintäänkin ylläpitävää harjoittelua voi tehdä omalla kehonpainolla yksin tai parin kanssa ja tarvittaessa erilaisia yksinkertaisia välineitä käyttäen.
Vastustuskyvyn kannalta olisi hyvä, jos harjoittelu ei olisi intensiteetiltään ja määrältään kehoa toistuvasti äärirajoille vievää nykyisessä epidemiassa.
Käy lukemassa vieraskirjoitus Lihastohtorin blogista harjoittelemattomuusjakson vaikutuksesta lihaksen kokoon ja voimaan.
Artikkelin yhteenveto:
Alle neljän viikon harjoittelemattomuusjakso ei vaikuta voimatasoihin juuri ollenkaan varsinkin kokeneemmilla ja vanhemmilla treenaajilla. Tämän jälkeen voima alkaa vähitellen vähenemään.
Lihakset alkavat hiljalleen surkastua noin kolmen viikon treenitauon jälkeen.
Treenitauko saattaa liioitella lihasten pienentymistä. Älä säikähdä sitä, sillä tämä johtuu osittain lihasten glykogeenivarastojen pienentymisestä ja viimeisen treenin aiheuttaman pienen lihasturvotuksen katoamisesta, ei ”oikean” lihasmassan häviämisestä. Tämä vaikutus on väliaikainen, ja jo pienikin harjoittelu palauttaa glykogeenitasot lihaksessa.
Pidemmän harjoittelemattomuusjakson jälkeen jälleen aloitettu harjoittelu palauttaa menetetyn lihasmassan ja voiman takaisin hyvin nopeasti ”lihasmuistin” ansiosta.
Olen pääkaupunkiseudulla vaikuttava voima-, fysiikka- ja yleisurheiluvalmentaja. Valmentajana olen toiminut kohta 10 vuoden ajan. Olen erityisen kiinnostunut urheilijoiden suorituskyvystä ja harjoittelun yksilöllisistä eroista urheilijoilla sekä periodisaatiosta. Saan urheilijat suoriutumaan lajissaan paremmin.
Tämä sivusto on luotu aputyökaluksi valmentajille, jotka haluavat perustaa valmentamisensa tieteelliseen näyttöön. Alalla on vielä paljon käytössä vanhentuneita toimintatapoja, jotka heikentävät urheilijoiden suorituskykyä. Pyrin jakamaan tieteellistä tietoa ja auttamaan objektiivisesti kaikkia valmentajia.