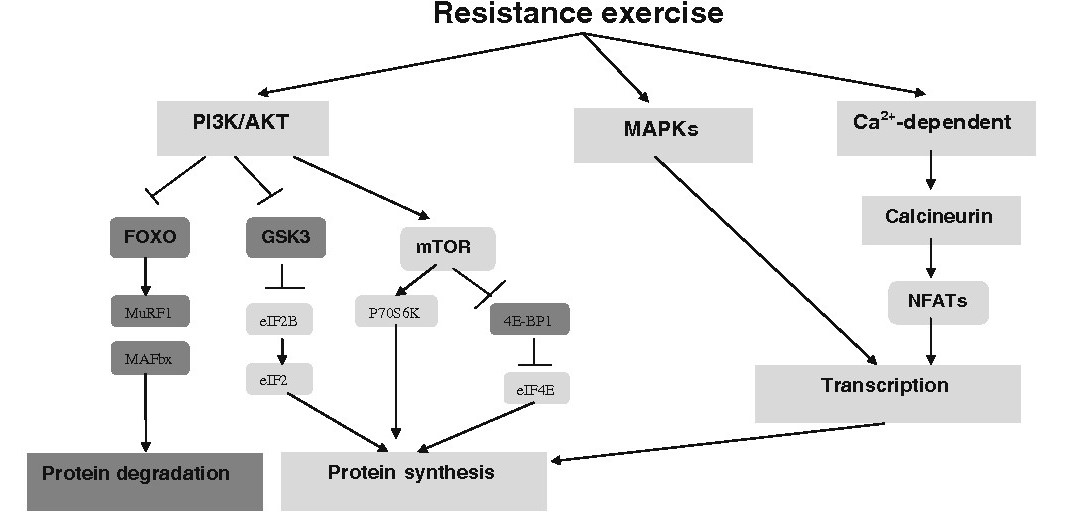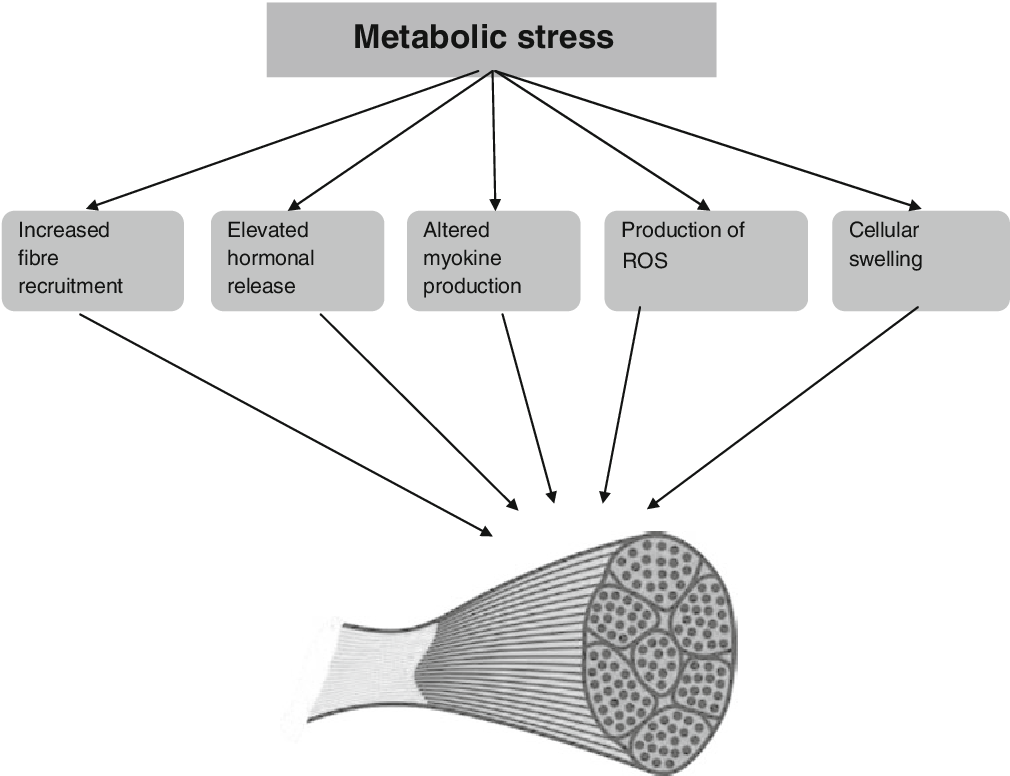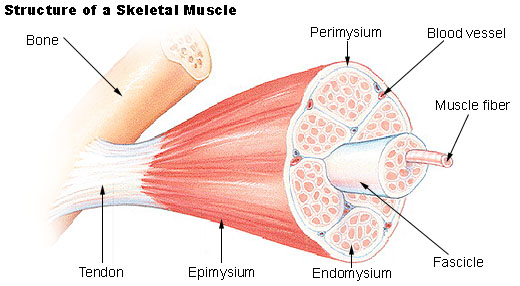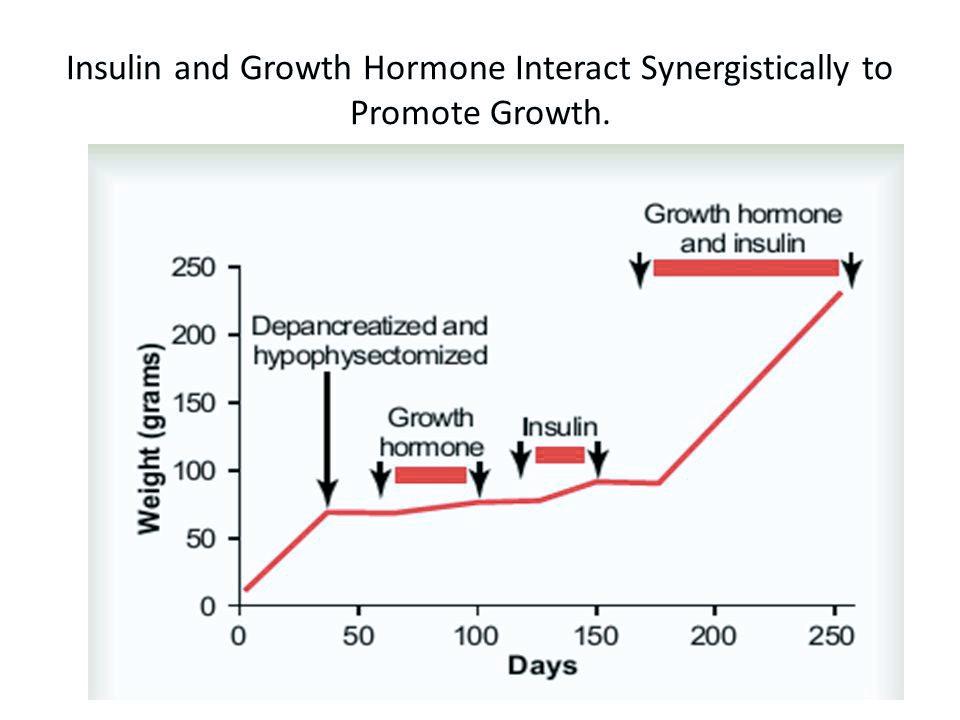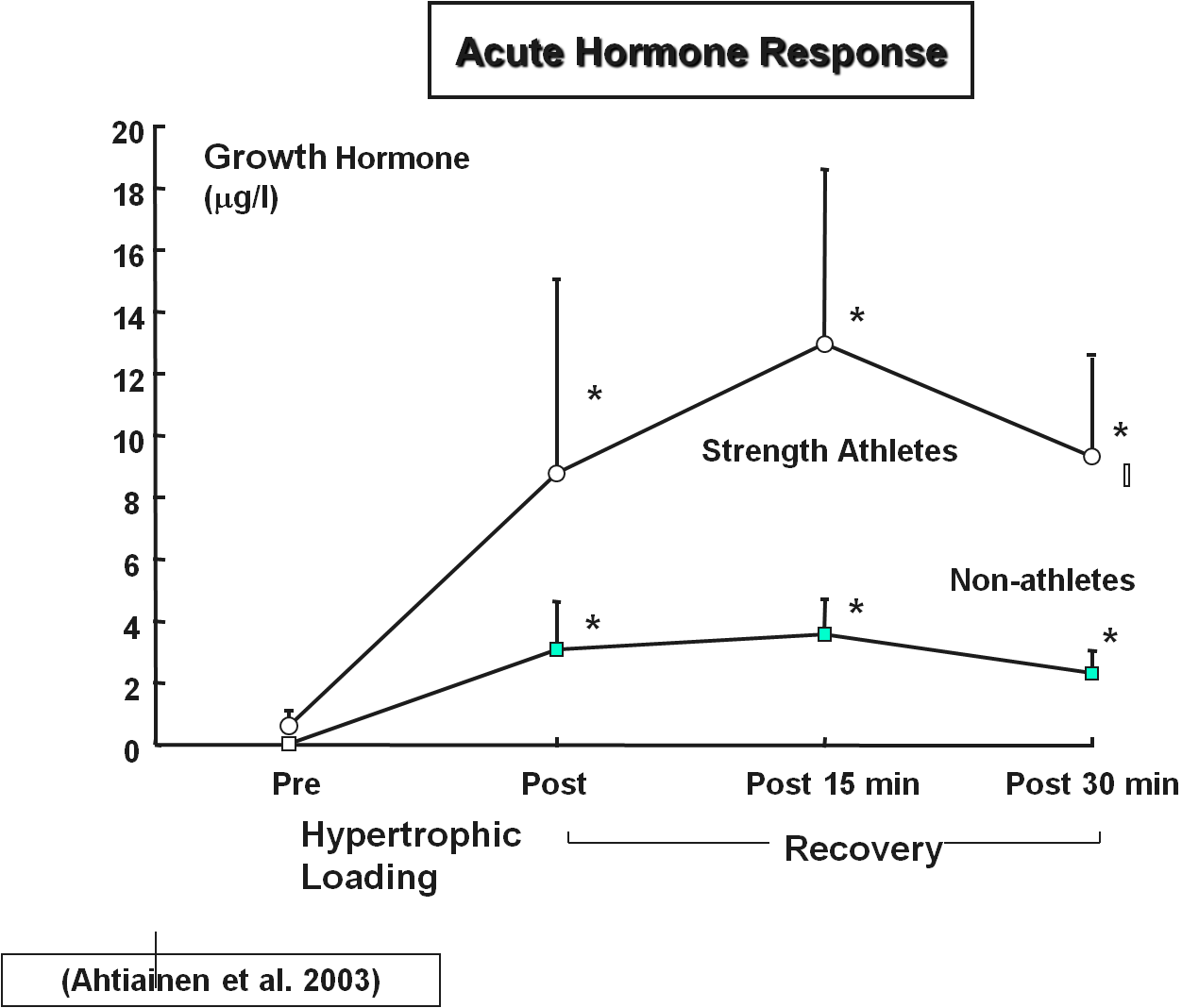
Lihahypertrofia osa 4/4: Yksilölliset vasteet voimaharjoitteluun ja harjoittelun periodisaatio
Lihaskasvua käsittelevä artikkelisarja lähenee loppua ja on viimeisen osan vuoro. Tässä artikkelissa käsitellään miten eri yksilöt adaptoituvat voimaharjoitteluun ja miten harjoittelu kannattaisi ohjelmoida, jotta kehitys olisi kaikkein optimaalisinta.
Genetiikan vaikutus lihaskasvuun
Geenit vaikuttavat suuresti henkilön kehittymiseen. Teoreettisesti voidaan ajatella maksimaalisen lihaskasvun rajojen olevan olemassa. Nämä määrittyvät genotyypin, eli geneettisen informaation, ja näiden geenien ilmentymisen yksilössä mukaan. Geenit asettavat teoreettisesti meille tietyt reunaehdot, miten reagoimme erilaisiin tilanteisiin. Geenien takia jotkut yksilöt kehittyvät nopeammin ja jotkut yksilöt hitaammin. Kuiinka paljon vaihtelua voi sitten eri henkilöiden välillä olla?
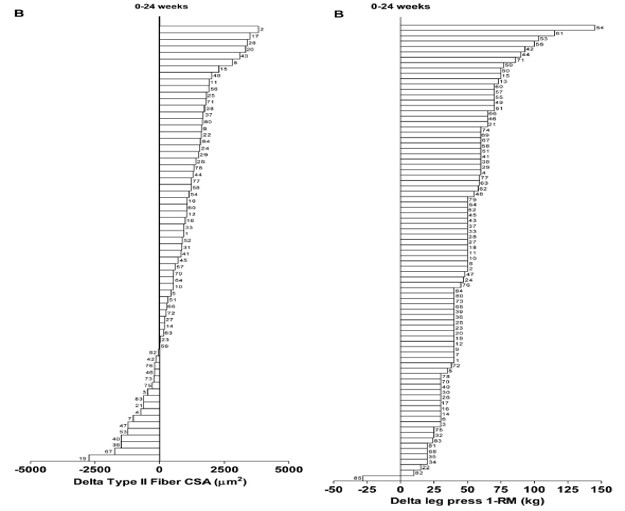
Yksilölliset vasteet harjoitteluun vaihtelevat hurjasti ihmisillä. Tämä tarkoittaa sitä, että jos kaksi samaa ihmistä tekee täysin saman ohjelman niin tuskin koskaan he kehittyvät täsmälleen saman verran. Esimerkiksi samanlaisella harjoittelulla lihaksen kasvu voi vaihdella jopa -11 % - + 30 % välillä ja voima taas - 8 % - + 60 % välillä (Ahtiainen et al., 2016). Ei-respondoivia ihmisiä, eli niitä, jotka eivät reagoi mitenkään tutkimusten voimaharjoitteluun on keskimäärin 25% koehenkilöistä (Bamman et al., 2007). Toisaalta nykyään ajatellaan, että erilainen harjoittelu, ravinto, lepo tai joku muu muuttuja olisi voinut vaikuttaa yksilöllisiin vasteisiin. Eli tälläiset ei-respondoivat koehenkilöt johtuvat pääosin huonosta tutkimusasetelmasta, elämäntilanteesta tai muusta tekijästä, mikä estää kehityksen. Jotkut ei-respondoivista ihmisistä tarvitsevat vain enemmän ja pidempää harjoittelua verrattuna toisiin.
Mikä näitä eroja sitten selittää? Pescatello ja kummanit (2013) pystyivät tunnistamaan 17 geeniä, joiden uskotaan selittävän osan henkilöiden välisistä eroista. Näistä tärkeimpänä hypertrofian kannalta pidetään tällä hetkellä BMP2-geeniä (Devaney et al., 2009). Ja myös MGF:ää pidetään erittäin tärkeänä (Bamman et al., 2007).
Lisäksi satelliittisolut lisääntyvät ja luovuttavat tumia tehokkaammin hyvin respondoituvilla ihmisillä kuin verrataan ei-respondoituviin ihmisiin (Petrella et al., 2008). Myös lihaksen morfologia vaikuttaa suuresti lihaskasvuun. Mitä enemmän lihassoluja henkilöllä on, niin sen helpommin lihas kasvaa. Lihassolujen määrä on aika vakio ja hyperplasiaa (lihassolujen lisääntymistä) ei tapahdu enää 24 viikon jälkeen syntymästä. Lihassolujen määrä onkin pitkälti geenien määrittämä.
Jos vertaillaan mieskehonrakentajia ja tavallisia kadun tallaajia niin kehonrakentajilla on enemmän lihassoluja hauiksissa ja nämä solut olivat myös isompia verrattuna tavallisiin tallaajiin (MacDougall et al., 1984). Joten niillä, jotka päätyivät kehonrakentajiksi oli jo enemmän lihassoluja kuin muilla normaaleilla ihmisillä. Tämä on yksi syy, miksi kehonrakentajat reagoivat paremmin voimaharjoitteluun.
Nopeiden lihassolujen määrä kasvatettavassa lihaksessa on tärkeä tekijä sen suhteen miten voimaharjoittelu vaikuttaa lihakseen. Tyypin II - lihassolut kasvavat noin 50% enemmän verrattua tyypin I -lihassoluihin. Toisaalta tässäkin suhteessa on hurjasti henkilöiden välistä vaihtelua (Kosek et al., 2006). Tutkimuksia, jotka selittävät fysiologisia mekanismeja yksilöllisessä voimaharjoittelussa on saatavilla erittäin niukasti, mutta erot johtuvat yleisesti perimästä, geenien ilmenemisessä, soluviestinnässä, hormonireseptoreiden toiminnassa ja satelliittisoluissa. Nostan vielä esille ravitsemuksen, jota harvoin tutkimuksissa kontrolloidaan ja sillä on valtava merkitys lihaskasvun mahdollistamisessa.
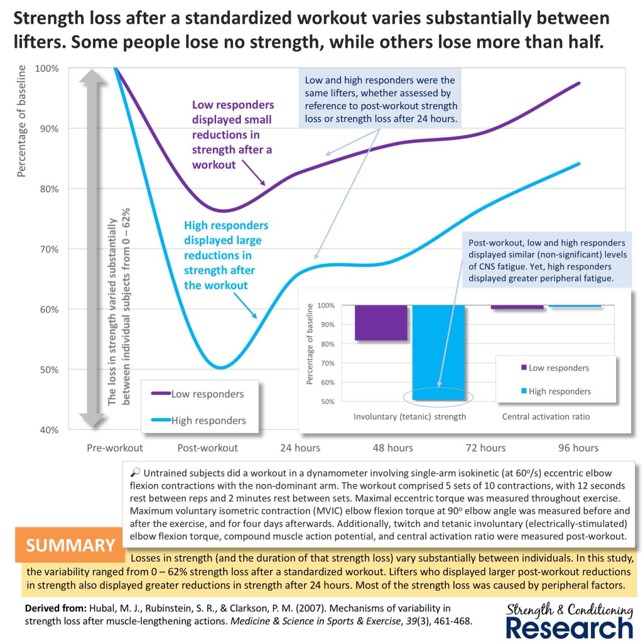
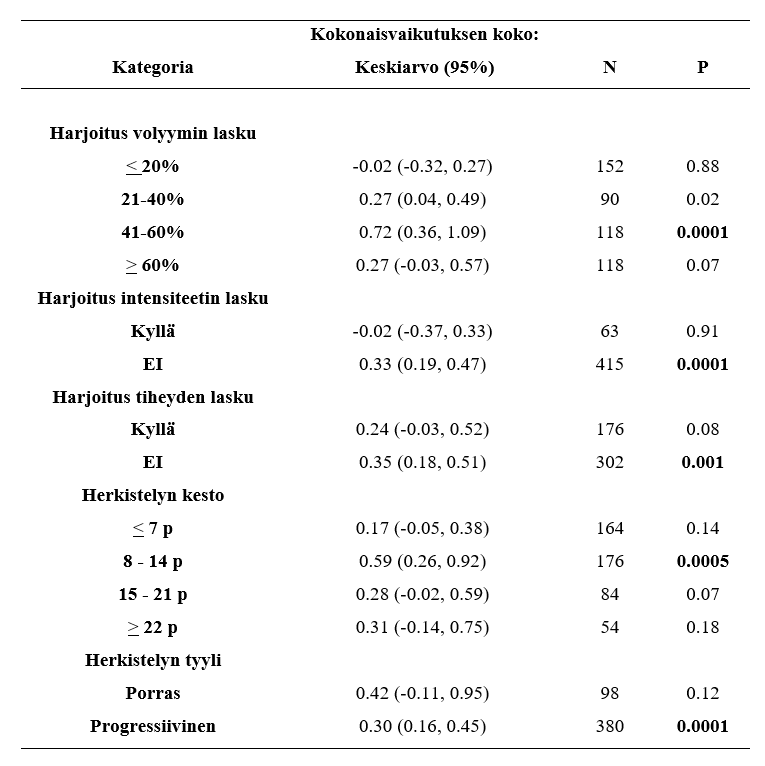
Genetiikalla on siis kieltämättä iso rooli henkilön lihaskasvussa. Voit kiittää vanhempiasi näistä lahjoista, mutta muuten asiaan ei kannata kiinnittää sen enempää huomiota. Jokainen voi ja pystyy kehittymään, vaikka tutkimuksissa usein löydetään ei-respontoivia henkilöitä harjoitteluun niin tähän saattaa olla syynä vääränlainen tutkimuksenasettelu, harjoittelu tai ohjelmointi. Yksilöllinen harjoittelu ja ohjelma mahdollistavat jokaisen kehittymisen. Heikosti respondoivien pitää joskus reenata jopa paljon vähemmän ja harvemmin kehittyäkseen. Tai kevyemmillä kuormilla. Myös harjoitustausta vaikuttaa suuresti henkilön kykyyn adaptoitua. Aloittelevilla ihmisillä lihaskasvu voi olla jopa 28% ja korkeimmillaan 58% 16 viikossa (Stewart & Rittweger, 2006), kun taas kehonrakentajat kehittyivät viidessä kuukaudessa “vain” 3-7% (Alway et al., 1992). Ohjelmointi ja ohjelman muuttujien manipulointi korostuu erityisesti, kun harjoitustaustaa kertyy enemmän. Kevennetyt jaksot voivat auttaa taas respontoimaan paremmin stimulukseen (Ogasawara et al., 2013).
Ei-respondereilla ei siis ole mielenvika pääsään vaan se on ihan oikea asia. Heidän tulee erityisesti keskittyä ravintoon ja oikeanlaiseen harjoitteluun. Huonosti respondoivat saattavat olla vain hitaammin respondoivia ja tarvitsevat enemmän harjoitusvolyymia kehittyäkseen. Nimi on vähän harhaanjohtava sillä jokainen kehittyy oikeanlaisella harjoittelulla. Saatat vain olla heikommin ja hitaammin respondoituva henkilö verrattuna salikaveriisi. Lisäksi vielä useissa tutkimuksissa kehitys on 12 viikossa esim. non respondereilla “vain” 5%. Tämä tulos luokitellaan tutkimuksissa hitaasti reaspondoivaksi, mutta jos kehitys jatkuisi koko vuoden verran samanlaisena se vastaisi alle vuodessa esimerkiksi 100kg penkissä siirtymistä 121,6 kiloon. Kuinka moni olisi tyytyväinen tuohon kehitykseen vuodessa? Minä ainakin olisin. Joten nimi on hiukan harhaanjohtava ja yleisesti ongelmat johtuvat tutkimusasetelmasta, sekä heterogeenisestä tutkimusjoukosta.
Periodisaatio lihaskasvussa
Periodisaatiota on tutkittu antiikin ajoista saakka. Mutta 1950-luvulla Neuvostoliitossa keskityttiin erityisesti periodisaation tutkimiseen. Tuolloin kehitelty superkompensaatioajatus antoi perustan nykyiselle periodisaatiolle. Tuolloin kehitettiin perinteinen lineaarinen periodisaatio malli olympiakisoihin valmistautuessa (Stone et al., 2000).
Neuvostoliitossa periodisaation (ja ylipäätään liikuntatieteen) tutkimus- ja kehitystyö oli tuolloin huomattavan laajaa ja systemaattista. Tämän tutkimustyön tuloksena syntyi klassinen periodisaatiomalli. Muissakin maissa toki käytettiin ja kehitettiin samantyylistä periodisaatiota, tosin pienemmässä mittakaavassa.
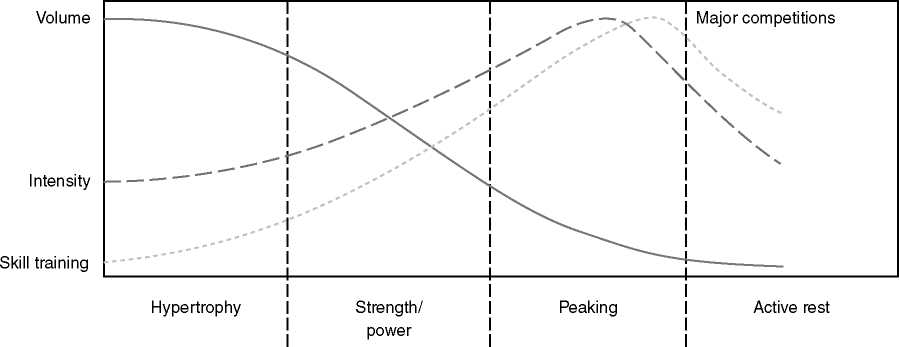
Periodisaatio on erittäin vähän tutkittu aihealue. Johtuen luultavasti siitä, että periodisaatio ei ole mikään selkeästi käsitettävä kokonaisuus. Erillaisia malleja voidaan esittää, mutta näidenkin sisällä voi olla loputtamattomasti erillaisia muutujia, joita voi siirrellä ja muutella. Periodisaatio perustuu Selyen yleiseeen adaptaatio syndrooma - teoriaan (GAS - general adaptation syndrome). Se perustuu kehon reaktioihin, jotka ovat hälytys, vastustus ja uupuminen. (Selye, 1950). Teoriaa on nykypäivinä kritisoitu yksinkertaistamisesta ja siitä, että se ei huomioi esimerkiksi psykologisia, teknisiä ja ravinnollisia seikkoja. Selyen teorian mukaan harjoittelua pidetään stressitekijänä, johon voidaan soveltaa GAS - teoriaa. Harjoittelun tulee olla progressiivisesti haastavaa, jotta tasannevaihetta ei tule harjoittelussa ja toisaalta harjoittelu ei saa olla ylikuluttavaa.
Periodisaatiomalleista kolmea on tutkittu lihashypertrofian kannalta: perinteinen lineaarinen malli, ei-lineaarinen ja käänteinen periodisaatio.
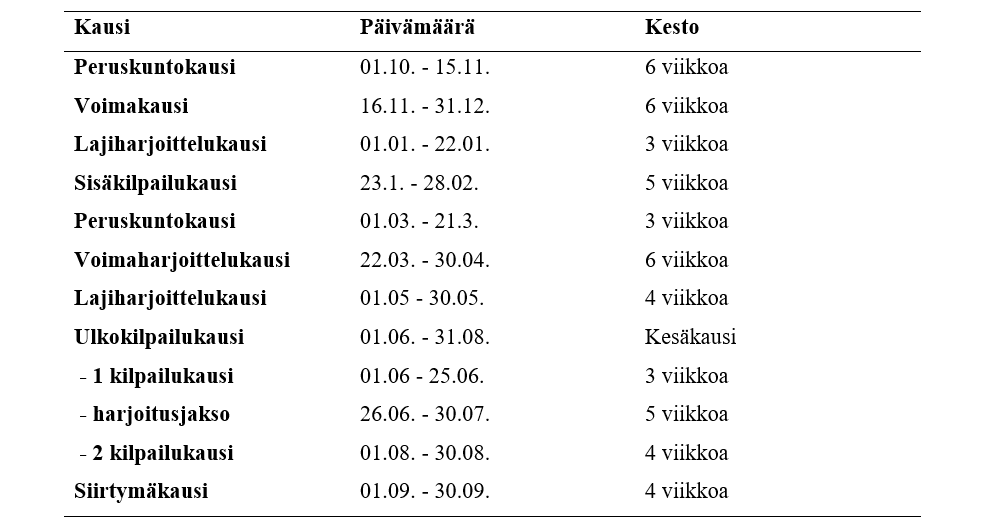
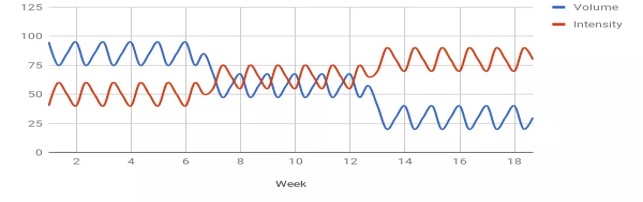
Perinteisessä lineaarisessa mallissa on vähintään makrosykli, mesosykli ja mikrosykli (esimerkiksi vuosi, peruskuntokausi 8 vko, yksi harjoitusviikko). Malli etenee korkeasta voluumista ja matalasta intensiteetistä kohti korkeaan intensiteettiin ja matalaan voluumiin. Esimerkiksi rakenteellinen jakso/hypertrofinen jakso, jossa voluumi nousee ja intensiteetti pysyy 60-75% välillä, seuraavaksi voi olla perusvoimakausi intensiteetin ollessa 80-90% välillä ja voluumin vähetessä. Viimeinen mesosykli olisi intensiteetiltään suurin 90-100%, mutta voluumiltaan pienin.
Lihaskasvun osalta aihetta on tutkittu jonkin verran, mutta itse pidän tutkimuksia metodologisilta osuuksilta ontuvilta, joten johtopäätöksiä ei tule vetää vielä näiden tutkimusten pohjalta. Stone ja kumppanit (1981) tutkivat periodisaatioidun ohjelman ja ei-periodisoidun ohjelman eroja, ja huomasivat hypertrofian (mitattu veden alaispunnituksella) eroavan tilastollisesti merkitsevästi ei-periodisoidusta ohjelmoinnista. Baker:in tutkimusporukka taa (1994) ei löytänyt eroja periodisatioidun ja ei-periodisaatiouidun välille (ihopoimumittaus). Kun taas Monteiro ja kumppanit (2009) löysivät eron (ihopoimu, ei tilastollisesti merkittävä) periodisoidun harjoittelun hyväksi.
Voiman osalta periodisoitu harjoittelu tuottaa kiistatta parempia tuloksia kuin ei periodisoitu harjoittelu (Ahmadizad et al., 2014;, Monteiro et al., 2009,Willoughby, 1993). Voisi ajatellakin, että koska voima nousee paremmin periodisoidulla harjoittelulla, johtaisi se mekaanisen kuormituksen johdosta myös suurempiin hypertrofisiin adaptaatioihin ajan kanssa, mutta tätä ei ole vielä todistettu.
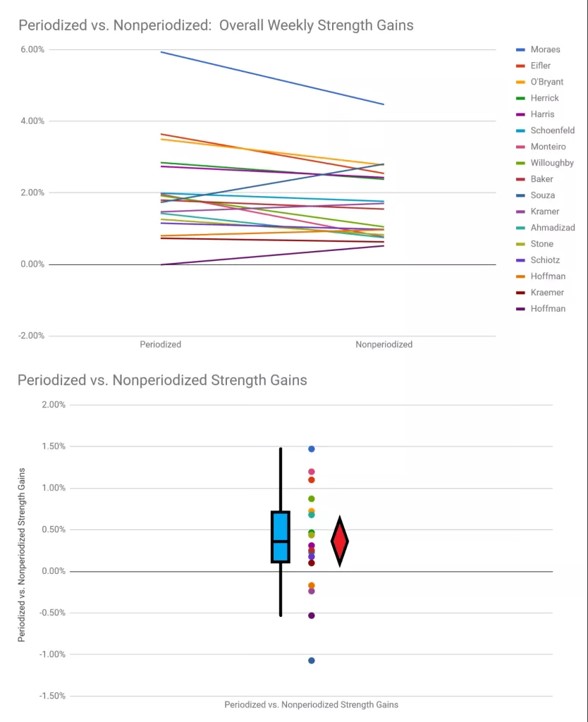
Ei-lineaarinen periodisaatio tai aaltoileva periodisaatio on nykyään yksi suosituimmista suuntauksista. Sitä on käytetty jo kauan, mutta Poliquin (1988) oli ensimmäinen, joka esitteli sen tiedeyhteisössä. Hänen mukaansa lineaarisen ohjelmoinnin ongelmat ovat liian suuri progressiivinen painon lisäys liian lyhyessä ajassa ja alussa saavutettujen hypertrofisten adaptaatioiden menetys muissa mesosykleissä, koska voluumi laskee liikaa.
Aaltoilevassa periodisaatiossa vaihdellaan voluumia ja intensiteettiä kuinkas muutenkaan kuin aaltoilevaan tyyliin. Mallia on kehitetty eteenpäin ja tällä hetkellä suosituin malli taitaa olla päivittäin vaihtuva periodisaatio (daily undulating periodization - DUP). Tässä mallissa on viikon sisällä maksimivoima, perusvoima ja hypertrofinen harjoitus. Aihetta on tutkittu suhteellisen laajasti. Ja vain yksi tutkimus sai tilastollisesti merkitsevän eron lineaarisen ja ei-lineaarisen mallin välille. Tässä tutkimuksessa ei-lineaarinen malli oli selvästi parempi lisäämään kyynärpään koukistajien ja ojentajien tiheyttä (Simao et al., 2012). Muuten muissa tutkimuksissa ei ole saatu eroja mallien välille lihaskasvun osalta, mikä osoittaa kummankin mallin olevan yhtä toimiva lihaskasvun osalta (Baker et al., 1994; Davitt et al., 2014; , Harries et al., 2015; Monteiro et al., 2009, Prestes et al., 2009, Simao et al., 2012). Toisaalta tutkimusten kesto on ollut korkeimmillaan vain 12 viikkoa ja kun puhutaan periodisaatiosta yleisesti urheilijoiden osalta puhutaan vuosista.
Itse suosin valmennettavillani enemmän hybridimallia, joka on versio aaltoilevasta mallista. Tässä mallissa harjoitusohjelma on laadittu blokkeihin ja näiden blokkien sisällä tapahtuu aaltoilua. Blokit toisaalta etenevät lineaarisesti. Viikon sisällä siinä tehdään maksimivoimasarjoja ja perusvoima sarjoja samassa harjoituksessa järkevästi ohjelmoiden. Pitkillä perusvoimakausilla saattaa hävitä liikaa maksimivoimaa ja pitkillä maksimivoima kausilla saattaa hävitä liikaa lihasta tai perusvoimaa, joten järkevä hybridiohjelmointi poistaa nämä ongelmat.
Käänteistä periodisaatiota on ehdotettu lihaskasvuun tähtääville tärkeimmäksi ohjelmaksi. Lineaarinen malli tiputtaa jatkuvasti voluumia kohti kauden päätähtäintä ja kun tiedetään voluumin olevan elintärkeä hypertrofian kannalta voisi ajatella lineaarisen mallin olevan huono lihaskasvun kannalta. Käänteisessä mallissa voluuumi on korkeimmillaan kauden päätähtäimen kohdalla. Aihe on vielä tutkimuskentällä tuore ja vain tietääkseni Prestes ja kumppanit (2009) ovat sitä tutkineet huonoin tuloksin käänteisen mallin puolesta. Lineaarinen malli oli huomattavasti parempi kuin käänteinen malli. Toisaalta mittauksissa käytettiin ihopoimupihtejä, jotka ovat metodisesti yhtä tyhjän kanssa.
Tiettyä suunnitelmallisuutta harjoitteluun tarvitaan, koska muuten harjoittelusta puuttuu variaatio, progressiivisuus ja tavoitteet. Lihaskasvun osalta yksi tärkeimmistä periodisaation tuomista asioista on kevennetyt jaksot. Keho ottaa paremmin vastaan harjoittelua kuin välillä tiputtaa voluumia ja antaa kropan palautua. Lisäksi on tärkeä kerätä palautetta periodisaation toimivuudesta ja suunnitelman muokkaaminen sen mukaan auttavat kehittämään ohjelmointia.
Kiitos lukijalle, jos jaksoit lukea tänne saakka! Alla lähdeluettelo kaikista artikkelisarjan osista. Aiheeseen liittyvä lukeminen ei ihan heti lopu kesken. Jätä palautetta tai kommentteja ja tsemppiä treeneihin!
Lähteet:
Ahtiainen, J., Pakarinen, A., Kraemer, W. & Häkkinen, K. (2003). Acute Hormonal and Neuromuscular Responses and Recovery to Forced vs. Maximum Repetitions Multiple Resistance Exercises. International Journal of Sports Medicine, 24(6), 410–418. doi: 10.1055/s-2003-41171
Ahtiainen, J. P., Pakarinen, A., Alen, M., Kraemer, W. J., & Häkkinen, K. (2005). Short vs. Long Rest Period Between the Sets in Hypertrophic Resistance Training: Influence on Muscle Strength, Size, and Hormonal Adaptations in Trained Men. The Journal of Strength and Conditioning Research, 19(3), 572. doi: 10.1519/15604.1
Ahtiainen JP, Walker S, Peltonen H, Holviala J, Sillanpää E, Karavirta L, et al. Heterogeneity in resistance training-induced muscle strength and mass responses in men and women of different ages. Age (Omaha). 2016;38:1–13.
Ahmadizad, S., Ghorbani, S., Ghasemikaram, M. & Bahmanzadeh, M. (2014) Effects of short-term nonperiodized, linear periodized and daily undulating periodized resistance training on plasma adiponectin, leptin and insulin resistance. Clinical biochemistry, 47: 417-422.
Alegre, L., Jimenez, F., Gonzalo-Orden, J., Acero, R. & Aguado, X. (2006). Effects of dynamic resistance training on fascicle length and isometric strength. Journal of sports sciences: 24. 501-8. 10.1080/02640410500189322.
Allen, D. G., Whitehead, N. P., & Yeung, E. W. (2005). Mechanisms of stretch-induced muscle damage in normal and dystrophic muscle: role of ionic changes. The Journal of Physiology, 567(3), 723–735. doi: 10.1113/jphysiol.2005.091694
Alway, S. E., Grumbt, W. H., Stray-Gundersen, J., & Gonyea, W. J. (1992). Effects of resistance training on elbow flexors of highly competitive bodybuilders. Journal of Applied Physiology, 72(4), 1512–1521. doi: 10.1152/jappl.1992.72.4.1512
Atherton, P. J., Babraj, J., Smith, K., Singh, J., Rennie, M. J., & Wackerhage, H. (2005). Selective activation of AMPK-PGC-1α or PKB-TSC2-mTOR signaling can explain specific adaptive responses to endurance or resistance training-like electrical muscle stimulation. The FASEB Journal, 19(7), 786–788. doi: 10.1096/fj.04-2179fje
Baker, D., Wilson, G. & Carolyn, R. (1994) Periodization: The effect on strength of manipulating volume and intensity. Journal of strength and conditioning research, 8: 235-242.
Bamman, M. M., Petrella, J. K., Kim, J.-S., Mayhew, D. L., & Cross, J. M. (2007). Cluster analysis tests the importance of myogenic gene expression during myofiber hypertrophy in humans. Journal of Applied Physiology, 102(6), 2232–2239. doi: 10.1152/japplphysiol.00024.2007
Benziane, B., Burton, T. J., Scanlan, B., Galuska, D., Canny, B. J., Chibalin, A. V., … Stepto, N. K. (2008). Divergent cell signaling after short-term intensified endurance training in human skeletal muscle. American Journal of Physiology-Endocrinology and Metabolism, 295(6). doi: 10.1152/ajpendo.90428.2008
Blazevich, A. J., Cannavan, D., Coleman, D. R., & Horne, S. (2007). Influence of concentric and eccentric resistance training on architectural adaptation in human quadriceps muscles. Journal of Applied Physiology, 103(5), 1565–1575. doi: 10.1152/japplphysiol.00578.2007
Bondesen, B. A., Mills, S. T., & Pavlath, G. K. (2006). The COX-2 pathway regulates growth of atrophied muscle via multiple mechanisms. American Journal of Physiology-Cell Physiology, 290(6). doi: 10.1152/ajpcell.00518.2005
Bondesen, B. A., Mills, S. T., Kegley, K. M., & Pavlath, G. K. (2004). The COX-2 pathway is essential during early stages of skeletal muscle regeneration. American Journal of Physiology-Cell Physiology, 287(2). doi: 10.1152/ajpcell.00088.2004
Brentano, M. & Martins Kruel, L. (2011) A review on strength exercise-induced muscle damage: applications, adaption mechanisms and limitations. J Sports Med Phys Fitness, 51(1): 1-10.
Burd, N. A., Holwerda, A. M., Selby, K. C., West, D. W. D., Staples, A. W., Cain, N. E., … Phillips, S. M. (2010). Resistance exercise volume affects myofibrillar protein synthesis and anabolic signalling molecule phosphorylation in young men. The Journal of Physiology, 588(16), 3119–3130. doi: 10.1113/jphysiol.2010.192856
Burd, N. A., Mitchell, C. J., Churchward-Venne, T. A., & Phillips, S. M. (2012). Bigger weights may not beget bigger muscles: evidence from acute muscle protein synthetic responses after resistance exercise. Applied Physiology, Nutrition, and Metabolism, 37(3), 551–554. doi: 10.1139/h2012-022
Choi J., Takahashi H., Itai Y. (1998) The difference between effects of ‘power-up type’ and ‘bulk-up type’ strength training exercises: With special reference to muscle cross-sectional area. Jpn J Phys Fitness Sports Med 47(1), 119-129.
Clarkson, P. M., Byrnes, W. C., Mccormick, K. M., Turcotte, L. P., & White, J. S. (1985). Muscle Soreness And Serum Creatine Kinase Activity Following Isometric, Eccentric And Concentric Exercise. Medicine & Science in Sports & Exercise, 17(2), 277. doi: 10.1249/00005768-198504000-00424
Coffey, V. G., Zhong, Z., Shield, A., Canny, B. J., Chibalin, A. V., Zierath, J. R., & Hawley, J. A. (2006). Early signaling responses to divergent exercise stimuli in skeletal muscle from well-trained humans. The FASEB Journal, 20(1), 190–192. doi: 10.1096/fj.05-4809fje
Cornelison, D. & Wold, B. (1997) Single-cell analysis of regulatory gene expression in quiescent and activated mouse skeletal muscle satellite cells. Advances in developmental biology, 191(2): 270–283.
Conboy, I. M., Conboy, M. J., Wagers, A. J., Girma, E. R., Weissman, I. L., & Rando, T. A. (2005). Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature, 433(7027): 760–764. doi: 10.1038/nature03260
Dangott, B., Schultz, E., & Mozdziak, P. E. (2000). Dietary Creatine Monohydrate Supplementation Increases Satellite Cell Mitotic Activity During Compensatory Hypertrophy. International Journal of Sports Medicine, 21(1), 13–16. doi: 10.1055/s-2000-8848
Davitt, P. M., Pellegrino, J. K., Schanzer, J. R., Tjionas, H., & Arent, S. M. (2014). The Effects of a Combined Resistance Training and Endurance Exercise Program in Inactive College Female Subjects. Journal of Strength and Conditioning Research, 28(7), 1937–1945. doi: 10.1519/jsc.0000000000000355
Devaney, J. M., Tosi, L. L., Fritz, D. T., Gordish-Dressman, H. A., Jiang, S., Orkunoglu-Suer, F. E., … Rogers, M. B. (2009). Differences in fat and muscle mass associated with a functional human polymorphism in a post-transcriptionalBMP2gene regulatory element. Journal of Cellular Biochemistry, 107(6), 1073–1082. doi: 10.1002/jcb.22209
Dhawan, J., & Rando, T. A. (2005). Stem cells in postnatal myogenesis: molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends in Cell Biology, 15(12), 666–673. doi: 10.1016/j.tcb.2005.10.007
Egner, I. M., Bruusgaard, J. C., Eftestøl, E., & Gundersen, K. (2013). A cellular memory mechanism aids overload hypertrophy in muscle long after an episodic exposure to anabolic steroids. The Journal of Physiology, 591(24): 6221–6230. doi: 10.1113/jphysiol.2013.264457
Evans, W. J., & Cannon, J. G. (1991). The Metabolic Effects of Exercise-Induced Muscle Damage. Exercise and Sport Sciences Reviews, 19(1). doi: 10.1249/00003677-199101000-00003
Finkenzeller, G., Newsome, W., Lang, F., & Häussinger, D. (1994). Increase of c-jun mRNA upon hypo-osmotic cell swelling of rat hepatoma cells. FEBS Letters, 340(3), 163–166. doi: 10.1016/0014-5793(94)80129-0
Folland, J. & Williams, A. (2007) The adaptations to strength training - morphological and neurological contributions to increased strength. Sports medicine, 37 (2): 145-168.
Fonseca, R. M., Roschel, H., Tricoli, V., Souza, E. O. D., Wilson, J. M., Laurentino, G. C., … Ugrinowitsch, C. (2014). Changes in Exercises Are More Effective Than in Loading Schemes to Improve Muscle Strength. Journal of Strength and Conditioning Research, 28(11), 3085–3092. doi: 10.1519/jsc.0000000000000539
Fry, C. S., Glynn, E. L., Drummond, M. J., Timmerman, K. L., Fujita, S., Abe, T., … Rasmussen, B. B. (2010). Blood flow restriction exercise stimulates mTORC1 signaling and muscle protein synthesis in older men. Journal of Applied Physiology, 108(5), 1199–1209. doi: 10.1152/japplphysiol.01266.2009
Gardiner NE. Athletics of the ancient world. Oxford: University Press; 1930.
Glass, D. J. (2010). PI3 Kinase Regulation of Skeletal Muscle Hypertrophy and Atrophy. Current Topics in Microbiology and Immunology Phosphoinositide 3-Kinase in Health and Disease, 267–278. doi: 10.1007/82_2010_78
Goodman, C. A., Mayhew, D. L., & Hornberger, T. A. (2011). Recent progress toward understanding the molecular mechanisms that regulate skeletal muscle mass. Cellular Signalling, 23(12), 1896–1906. doi: 10.1016/j.cellsig.2011.07.013
Gordon, S. E., Kraemer, W. J., Vos, N. H., Lynch, J. M., & Knuttgen, H. G. (1994). Effect of acid-base balance on the growth hormone response to acute high-intensity cycle exercise. Journal of Applied Physiology, 76(2), 821–829. doi: 10.1152/jappl.1994.76.2.821
Goto, K., Ishii, N., Kizuka, T. & Takamatsu, K. (2005) The impact of metabolic stress on hormonal responses and muscular adaptations. Medicine and science in sports and exercise, 37(6): 955-963.
Guyton. A. C., Hall J. E. 2011. Textbook of Medical Physiology. 12th edition. Philadelphia, USA : W.B. Saunders Company
Harries, S. K., Lubans, D. R., & Callister, R. (2016). Comparison of resistance training progression models on maximal strength in sub-elite adolescent rugby union players. Journal of Science and Medicine in Sport, 19(2), 163–169. doi: 10.1016/j.jsams.2015.01.007
Hackett, D. A., Johnson, N. A., & Chow, C.-M. (2013). Training Practices and Ergogenic Aids Used by Male Bodybuilders. Journal of Strength and Conditioning Research, 27(6), 1609–1617. doi: 10.1519/jsc.0b013e318271272a
Handayaningsih, A.-E., Iguchi, G., Fukuoka, H., Nishizawa, H., Takahashi, M., Yamamoto, M., … Takahashi, Y. (2011). Reactive Oxygen Species Play an Essential Role in IGF-I Signaling and IGF-I-Induced Myocyte Hypertrophy in C2C12 Myocytes. Endocrinology, 152(3), 912–921. doi: 10.1210/en.2010-0981
Harber, M. P., Konopka, A. R., Douglass, M. D., Minchev, K., Kaminsky, L. A., Trappe, T. A., & Trappe, S. (2009). Aerobic exercise training improves whole muscle and single myofiber size and function in older women. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 297(5). doi: 10.1152/ajpregu.00354.2009
Harber, M. P., Konopka, A. R., Undem, M. K., Hinkley, J. M., Minchev, K., Kaminsky, L. A., … Trappe, S. (2012). Aerobic exercise training induces skeletal muscle hypertrophy and age-dependent adaptations in myofiber function in young and older men. Journal of Applied Physiology, 113(9), 1495–1504. doi: 10.1152/japplphysiol.00786.2012
Harridge, S. (2007) Plasticity of human skeletal muscle: Gene expression to in vivo function. Exp Physiolo. 92: 738-797.
Helms, E., Aragon, A. & Fitschen, P. (2014). Evidence-based recommendations for natural bodybuilding contest preparation: Nutrition and supplementation. Journal of the International Society of Sports Nutrition. 11. 20. 10.1186/1550-2783-11-20.
Hill, M., Wernig, A., & Goldspink, G. (2003). Muscle satellite (stem) cell activation during local tissue injury and repair. Journal of Anatomy, 203(1), 89–99. doi: 10.1046/j.1469-7580.2003.00195.x
Hornberger, T. A., Chu, W. K., Mak, Y. W., Hsiung, J. W., Huang, S. A., & Chien, S. (2006). The role of phospholipase D and phosphatidic acid in the mechanical activation of mTOR signaling in skeletal muscle. Proceedings of the National Academy of Sciences, 103(12), 4741–4746. doi: 10.1073/pnas.0600678103
Izquierdo, M., Ibañez, J., González-Badillo, J. J., Häkkinen, K., Ratamess, N. A., Kraemer, W. J., French, D., Eslava, J., Altadill, A., Asiain X. & Gorostiaga, E. M. (2006). Differential effects of strength training leading to failure versus not to failure on hormonal responses, strength, and muscle power gains. Journal of Applied Physiology, 100(5), 1647–1656. doi: 10.1152/japplphysiol.01400.2005
Jacinto, E., & Hall, M. N. (2003). TOR signalling in bugs, brain and brawn. Nature Reviews Molecular Cell Biology, 4(2), 117–126. doi: 10.1038/nrm1018
Kadi, F., Eriksson, A., Holmner, S., Butler-Browne, G. & Thornell, L. (1999) Cellular adaptation of the trapezius muscle in strength-trained athletes. Histochemistry and cell biology, 111(3): 189-95.
Kadi, F. & Thornell, L.E. (2000) Concomitant increases in myonuclear and satellite cell content of female trapezius muscle following strength training. Histochemistry and cell biology 113, 99-103.
Keogh, J. W., Wilson, G. J., & Weatherby, R. E. (1999). A Cross-Sectional Comparison of Different Resistance Training Techniques in the Bench Press. Journal of Strength and Conditioning Research, 13(3), 247–258. doi: 10.1519/00124278-199908000-00012
Kohn, T. A., Essén-Gustavsson, B., & Myburgh, K. H. (2010). Specific muscle adaptations in type II fibers after high-intensity interval training of well-trained runners. Scandinavian Journal of Medicine & Science in Sports, 21(6), 765–772. doi: 10.1111/j.1600-0838.2010.01136.x
Kosek, D. J., Kim, J.-S., Petrella, J. K., Cross, J. M., & Bamman, M. M. (2006). Efficacy of 3 days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in young vs. older adults. Journal of Applied Physiology, 101(2), 531–544. doi: 10.1152/japplphysiol.01474.2005
Krieger, J. W. (2010). Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy: A Meta-Analysis. Journal of Strength and Conditioning Research, 24(4), 1150–1159. doi: 10.1519/jsc.0b013e3181d4d436
Lang, F (2007). Mechanisms and Significance of Cell Volume Regulation. J Am Coll Nutr., 26: 613s-623s.
Lang, F., Busch, G. L., Ritter, M., Völkl, H., Waldegger, S., Gulbins, E., & Häussinger, D. (1998). Functional Significance of Cell Volume Regulatory Mechanisms. Physiological Reviews, 78(1), 247–306. doi: 10.1152/physrev.1998.78.1.247
Loenneke, J. P., Wilson, J. M., Marín, P. J., Zourdos, M. C., & Bemben, M. G. (2011). Low intensity blood flow restriction training: a meta-analysis. European Journal of Applied Physiology, 112(5), 1849–1859. doi: 10.1007/s00421-011-2167-x
Loenneke, J. P., Thiebaud, R. S., & Abe, T. (2014). Does blood flow restriction result in skeletal muscle damage? A critical review of available evidence. Scandinavian Journal of Medicine & Science in Sports, 24(6). doi: 10.1111/sms.12210
Lynn, R. & Morgan, D. (1994) Decline running produces more sarcomeres in rat vastus intermedius muscle fibers than does incline running. Journal of applied physiology: 77: 1439-1444.
Malm, C. (2001). Exercise-induced muscle damage and inflammation: fact or fiction? Acta Physiologica Scandinavica, 171(3), 233–239. doi: 10.1046/j.1365-201x.2001.00825.x
Macdougall, J. D., Gibala, M., Tarnopolsky, M., Macdonald, J., Interisano, S., & Yarasheski, K. (1995). The Time Course For Elevated Muscle Protein Synthesis Following Heavy Resistance Exercise. Medicine & Science in Sports & Exercise, 27(Supplement). doi: 10.1249/00005768-199505001-00367
MacDougall, D. Ward, G., Sale, D. & Sutton, J. (1977) Biochemical adaptation of human skeletal muscle to heavy resistance training and immobilization. Journal of applied physiology: respiratory, environmental and exercise physiology: 43(4): 700-3.
Macdougall, J. D., Sale, D. G., Alway, S. E., & Sutton, J. R. (1984). Muscle fiber number in biceps brachii in bodybuilders and control subjects. Journal of Applied Physiology, 57(5), 1399–1403. doi: 10.1152/jappl.1984.57.5.1399
Mascher, H., Andersson, H., Nilsson, P.-A., Ekblom, B., & Blomstrand, E. (2007). Changes in signalling pathways regulating protein synthesis in human muscle in the recovery period after endurance exercise. Acta Physiologica, 191(1), 67–75. doi: 10.1111/j.1748-1716.2007.01712.x
Masuda K., Choi J.Y., Shimojo H., Katsuta S. (1999) Maintenance of myoglobin concentration in human skeletal muscle after heavy resistance training. European Journal of Applied Physiology and Occupational Physiology 79(4), 347-352. [PubMed] [Google Scholar]
Martineau, L. & Gardiner, P. (2002) Skeletal muscle is sensitive to the tension-time integral but not to the rate of change of tension, as assessed by mechanically induced signaling. Journal of biomechanic, 35: 657-663.
Mchugh, M. P. (2003). Recent advances in the understanding of the repeated bout effect: the protective effect against muscle damage from a single bout of eccentric exercise. Scandinavian Journal of Medicine and Science in Sports, 13(2), 88–97. doi: 10.1034/j.1600-0838.2003.02477.x
Mcpherron, A. C., Lawler, A. M., & Lee, S.-J. (1997). Regulation of skeletal muscle mass in mice by a new TGF-p superfamily member. Nature, 387(6628), 83–90. doi: 10.1038/387083a0
Mikkola, J., Rusko, H., Izquierdo, M., Gorostiaga, E., & Häkkinen, K. (2012). Neuromuscular and Cardiovascular Adaptations During Concurrent Strength and Endurance Training in Untrained Men. International Journal of Sports Medicine, 33(09), 702–710. doi: 10.1055/s-0031-1295475
Moore, D., Phillips, S., Babraj, J., Smith, K. & Rennie, M. (2005) Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. American journal of physiology - endocrinology andmetabolism, 288: E1153–E1159.
Monteiro, A., Aoki., M., Evangelista,A., Alveno, D., Monteiro, G., Picarro Ida, C. & Ugrinowitcch, C. (2009) Nonlinear periodization maximizes strength gains in split resistance training routines. Journal of strength and conditioning research, 23: 1321-1326.
Netreba, A., Popov, D., Bravyy, Y., Lyubaeva, E., Terada, M., Ohira, T., Okabe, H., Vinogradova, O.& Ohira, Y. (2013). Responses of knee extensor muscles to leg press training of various types in human. Rossiĭskii fiziologicheskiĭ zhurnal imeni I.M. Sechenova / Rossiĭskaia akademiia nauk. 99. 406-16.
Nguyen, H. X., & Tidball, J. G. (2003). Null Mutation of gp91phoxReduces Muscle Membrane Lysis During Muscle Inflammation in Mice. The Journal of Physiology, 553(3), 833–841. doi: 10.1113/jphysiol.2003.051912
Nosaka, K., Lavender, A., Newton, M. & Sacco, P. (2003) Muscle damage in resistance training – is muscle damage necessary for strength gain and muscle hypertrophy? International journal of sport and health science, 1(1):1-8.
Nuckols, G. (2018) Periodization: What the data say. https://www.strongerbyscience.com/periodization-data/
Ogasawara, R., Kobayashi, K., Tsutaki, A., Lee, K., Abe, T., Fujita, S., … Ishii, N. (2013). mTOR signaling response to resistance exercise is altered by chronic resistance training and detraining in skeletal muscle. Journal of Applied Physiology, 114(7), 934–940. doi: 10.1152/japplphysiol.01161.2012
Pescatello, L. S., Devaney, J. M., Hubal, M. J., Thompson, P. D., & Hoffman, E. P. (2013). Highlights from the Functional Single Nucleotide Polymorphisms Associated with Human Muscle Size and Strength or FAMuSS Study. BioMed Research International, 2013, 1–11. doi: 10.1155/2013/643575
Petrella, J. K., Kim, J.-S., Mayhew, D. L., Cross, J. M., & Bamman, M. M. (2008). Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition: a cluster analysis. Journal of Applied Physiology, 104(6), 1736–1742. doi: 10.1152/japplphysiol.01215.2007
Pinto, R., Gomes, N., Radaelli, R., Botton, C., Brown, L. & Bottaro, M. (2011). Effect of Range of Motion on Muscle Strength and Thickness. Journal of strength and conditioning research / National Strength & Conditioning Association. 26. 2140-5. 10.1519/JSC.0b013e31823a3b15.
Poliquin, C. (1988) Five steps to increasing the effectiveness of your strength training program. Journal of strength and conditioning association. 10: 34-39.
Prestes, J., Frollini, A. B., Lima, C. D., Donatto, F. F., Foschini, D., Marqueti, R. D. C., … Fleck, S. J. (2009). Comparison Between Linear and Daily Undulating Periodized Resistance Training to Increase Strength. Journal of Strength and Conditioning Research, 23(9), 2437–2442. doi: 10.1519/jsc.0b013e3181c03548
Qaisar, R., Bhaskaran, S. & Remmen, H. (2016). Muscle fiber type diversification during exercise and regeneration. Free Radical Biology and Medicine. 98. 10.1016/j.freeradbiomed.2016.03.025.
Radaelli, R. J Fleck, S., Leite, T., Leite, R., Pinto, R., Fernandesm L. Simão, R. (2014). Dose Response of 1, 3 and 5 Sets of Resistance Exercise on Strength, Local Muscular Endurance and Hypertrophy. The Journal of Strength and Conditioning Research. 29. 10.1519/JSC.0000000000000758.
Ratamess, N. A., Falvo, M. J., Mangine, G. T., Hoffman, J. R., Faigenbaum, A. D., & Kang, J. (2007). The effect of rest interval length on metabolic responses to the bench press exercise. European Journal of Applied Physiology, 100(1), 1–17. doi: 10.1007/s00421-007-0394-y
Schoenfeld, B. (2010) The mechanism of muscle hypertrophy and their application to resistance training. Journal of strength and conditioning research, 24: 2857-2872.
Schoenfeld, BJ. Science and Development of Muscle Hypertrophy. Champaign, IL; Human Kinetics, 2016.
Schoenfeld, B, Ogborn, D. & Krieger, J. (2017) Dose-response relationship between weekly resistance training volume and increases in muscle mass: A systematic review and meta-analysis, Journal of Sports Sciences, 35:11, 1073-1082, DOI: 10.1080/02640414.2016.1210197
Schoenfeld, B. J., Ratamess, N. A., Peterson, M. D., Contreras, B., Sonmez, G. T., & Alvar, B. A. (2014). Effects of Different Volume-Equated Resistance Training Loading Strategies on Muscular Adaptations in Well-Trained Men. Journal of Strength and Conditioning Research, 28(10), 2909–2918. doi: 10.1519/jsc.0000000000000480
Schoenfeld, B, Grgic, J., Ogbord, D. & Krieger, J. (2017) Strength and hypertrophy adaptations between low- versus high-load resistance training: A systematic review and meta-analysis. The Journal of strength and conditioning research, Ahead of Print DOI: 10.1519/JSC.0000000000002200
Schoenfeld, B., Contreras, B., Vigotsky, A., Ogborn, D., Fontana, F. & Tiryaki-Sonmez, R. (2016). Upper body muscle activation during low-versus high-load resistance exercise in the bench press. 24. 217-224. 10.3233/IES-160620.
Schoenfeld, B. J., Ratamess, N. A., Peterson, M. D., Contreras, B., & Tiryaki-Sonmez, G. (2015). Influence of Resistance Training Frequency on Muscular Adaptations in Well-Trained Men. Journal of Strength and Conditioning Research, 29(7), 1821–1829. doi: 10.1519/jsc.0000000000000970
Schoenfeld, B. J., Peterson, M. D., Ogborn, D., Contreras, B., & Sonmez, G. T. (2015). Effects of Low- vs. High-Load Resistance Training on Muscle Strength and Hypertrophy in Well-Trained Men. Journal of Strength and Conditioning Research, 29(10), 2954–2963. doi: 10.1519/jsc.0000000000000958
Schoenfeld, B. J., Ogborn, D. I., & Krieger, J. W. (2015). Effect of Repetition Duration During Resistance Training on Muscle Hypertrophy: A Systematic Review and Meta-Analysis. Sports Medicine, 45(4), 577–585. doi: 10.1007/s40279-015-0304-0
Schoenfeld, B. J., Contreras, B., Krieger, J., Grgic, J., Delcastillo, K., Belliard, R., & Alto, A. (2019). Resistance Training Volume Enhances Muscle Hypertrophy but Not Strength in Trained Men. Medicine & Science in Sports & Exercise, 51(1), 94–103. doi: 10.1249/mss.0000000000001764
Schoenfeld, B.J. & Grgic, J. (2020) Effects of range of motion on muscle development during resistance training interventions: A systematic review. SAGE open medicine, 8: 1-8. https://doi.org/10.1177/2050312120901559
Schuenke, M. D., Herman, J. R., Gliders, R. M., Hagerman, F. C., Hikida, R. S., Rana, S. R., … Staron, R. S. (2012). Early-phase muscular adaptations in response to slow-speed versus traditional resistance-training regimens. European Journal of Applied Physiology, 112(10), 3585–3595. doi: 10.1007/s00421-012-2339-3
Selye, H. (1950). Stress and the General Adaptation Syndrome. Bmj, 1(4667), 1383–1392. doi: 10.1136/bmj.1.4667.1383
Serrano, A. L., Baeza-Raja, B., Perdiguero, E., Jardí, M., & Muñoz-Cánoves, P. (2008). Interleukin-6 Is an Essential Regulator of Satellite Cell-Mediated Skeletal Muscle Hypertrophy. Cell Metabolism, 7(1), 33–44. doi: 10.1016/j.cmet.2007.11.011
Simão, R., Spineti, J., Salles, B. F. D., Matta, T., Fernandes, L., Fleck, S. J., … Strom-Olsen, H. E. (2012). Comparison Between Nonlinear and Linear Periodized Resistance Training. Journal of Strength and Conditioning Research, 26(5), 1389–1395. doi: 10.1519/jsc.0b013e318231a659
Snyder, B. J., & Leech, J. R. (2009). Voluntary Increase in Latissimus Dorsi Muscle Activity During the Lat Pull-Down Following Expert Instruction. Journal of Strength and Conditioning Research, 23(8), 2204–2209. doi: 10.1519/jsc.0b013e3181bb7213
Snyder, B. J., & Fry, W. R. (2012). Effect of Verbal Instruction on Muscle Activity During the Bench Press Exercise. Journal of Strength and Conditioning Research, 26(9), 2394–2400. doi: 10.1519/jsc.0b013e31823f8d11
Stewart, C. & Rittweger, J. (2006) Adaptive processes in skeletal muscle: molecular regulators and genetic influences. Journal of musculoskeletal and neuronal interactions: 6(1): 73-86.
Stone, M., O’Bryant, H. & Garhammer J. (1981) Hypothetical model for strength training. Journal of sports medicine and physical fitness, 21: 342-351.
Stone, M. H., Potteiger, J. A., Pierce, K. C., Proulx, C. M., Obryant, H. S., Johnson, R. L., & Stone, M. E. (2000). Comparison of the Effects of Three Different Weight-Training Programs on the One Repetition Maximum Squat. The Journal of Strength and Conditioning Research, 14(3), 332. doi: 10.1519/1533-4287(2000)014<0332:coteot>2.0.co;2
Takarada, Y., Takazawa, H., Sato, Y., Takebayashi, S., Tanaka, Y., & Ishii, N. (2000). Effects of resistance exercise combined with moderate vascular occlusion on muscular function in humans. Journal of Applied Physiology, 88(6), 2097–2106. doi: 10.1152/jappl.2000.88.6.2097
Takarada, Y., Nakamura, Y., Aruga, S., Onda, T., Miyazaki, S., & Ishii, N. (2000). Rapid increase in plasma growth hormone after low-intensity resistance exercise with vascular occlusion. Journal of Applied Physiology, 88(1), 61–65. doi: 10.1152/jappl.2000.88.1.61
Tee, J. C., Bosch, A. N., & Lambert, M. I. (2007). Metabolic Consequences of Exercise-Induced Muscle Damage. Sports Medicine, 37(10), 827–836. doi: 10.2165/00007256-200737100-00001
Terzis, G., Spengos, K., Mascher, H., Georgiadis, G., Manta, P. & Blomstrand, E. (2010). The degree of p70S6k and S6 phosphorylation in human skeletal muscle in response to resistance exercise depends on the training volume. European journal of applied physiology. 110. 835-43. 10.1007/s00421-010-1527-2.
Thomas, G., & Hall, M. N. (1997). TOR signalling and control of cell growth. Current Opinion in Cell Biology, 9(6), 782–787. doi: 10.1016/s0955-0674(97)80078-6
Uchiyama, S., Tsukamoto, H., Yoshimura, S., & Tamaki, T. (2006). Relationship between oxidative stress in muscle tissue and weight-lifting-induced muscle damage. Pflügers Archiv - European Journal of Physiology, 452(1), 109–116. doi: 10.1007/s00424-005-0012-y
Wernbom, M., Augustsson, J. & Thomee, R. (2007). The Influence of Frequency, Intensity, Volume and Mode of Strength Training on Whole Muscle Cross-Sectional Area in Humans. Sports medicine (Auckland, N.Z.). 37. 225-64. 10.2165/00007256-200737030-00004.
Willoughby, D. (1993) The effects of mesocycle-length weight training programs involving periodization an partially equated volumes on upper and lower body strength. Journal of strength and conditioning research, 7: 2-8.
Wilson, J. M., Lowery, R. P., Joy, J. M., Loenneke, J. P., & Naimo, M. A. (2013). Practical Blood Flow Restriction Training Increases Acute Determinants of Hypertrophy Without Increasing Indices of Muscle Damage. Journal of Strength and Conditioning Research, 27(11), 3068–3075. doi: 10.1519/jsc.0b013e31828a1ffa
Wilson, J. M., Marin, P. J., Rhea, M. R., Wilson, S. M., Loenneke, J. P., & Anderson, J. C. (2012). Concurrent Training. Journal of Strength and Conditioning Research, 26(8), 2293–2307. doi: 10.1519/jsc.0b013e31823a3e2d
Wulf, G. (2013). Attentional focus and motor learning: a review of 15 years. International Review of Sport and Exercise Psychology, 6(1), 77–104. doi: 10.1080/1750984x.2012.723728
Yang, S. Y., & Goldspink, G. (2002). Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in myoblast proliferation and differentiation. FEBS Letters, 522(1-3), 156–160. doi: 10.1016/s0014-5793(02)02918-6
Zanou, N., & Gailly, P. (2013). Skeletal muscle hypertrophy and regeneration: interplay between the myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs) pathways. Cellular and Molecular Life Sciences, 70(21), 4117–4130. doi: 10.1007/s00018-013-1330-4
Zanchi, N. E., & Lancha, A. H. (2007). Mechanical stimuli of skeletal muscle: implications on mTOR/p70s6k and protein synthesis. European Journal of Applied Physiology, 102(3), 253–263. doi: 10.1007/s00421-007-0588-3