Oletko tehnyt harjoitusohjelman ja huomannut ohjelman jälkeen, ettet kehittynytkään tai urheilijasi ei saavuttanutkaan haluttua kehitystä? Kaikki yksilöt kehittyvät eri tahtiin ja tiedämme, että yksilöiden välillä on paljon eroja kehittymistahdissa. Jotkut yksilöt eivät vastaa tietynlaiseen harjoitteluun ollenkaan, kun toiset taas kehittyvät vimmatusti. Lihasmassaharjoittelun yhteydessä käytetään kansankielisesti termistöä hardgaineri, kun kehitystä ei tule. Mitä tämmöisissä tapauksissa kannattaa tehdä, jotta kehitystä saadaan jatkossa aikaan vai onko hardgainer tuomittu loppuelämäksi olemaan kehittymättä?
Mitkä kaikki vaikuttavat kehittymiseen voimaharjoittelussa?
Vaikka voimaharjoittelu onkin erinomainen tapa kehittää lihasvoimaa ja -kestävyyttä, se ei kuitenkaan ole aina yhtä tehokas kaikille ihmisille. Kehittymiseen vaikuttavat monet eri tekijät, kuten ikä, sukupuoli, perimä, elämäntavat, harjoitustausta ja mahdolliset sairaudet tai vammat.
Tämän vuoksi onkin tärkeää ymmärtää ja suunnitella voimaharjoittelua yksilöllisten tarpeiden mukaan. Yksilöllisyyden huomioiminen voimaharjoittelussa auttaa saavuttamaan parempia tuloksia ja välttämään mahdollisia vammoja tai ylirasitusta.
Erittäin tärkeä tekijä kehittymisessä on harjoitustausta. Esimerkiksi vasta-alkaja kehittyy huomattavia määriä alussa lihasmassan ja voiman osalta, kun taas kokeneemmalla harjoittelijalla pienenkin kehityksen saavuttaminen on haastavampaa (Rhea et al., 2003).
Lihaskasvusta ja kaikesta harjoittelusta puhuttaessa ei voi olla puhumatta ravitsemuksesta ja proteiinin merkityksestä. Yksilöt, jotka saavat riittävästi energiaa ja proteiinia tukemaan heidän harjoitteluaan, kehittyvät huomattavasti enemmän kuin ne, jotka eivät saa tarvitsemaansa ravintoa (Phillips and Van Loon, 2011). Voiko syyttää vain geenejään vai onko taustalla jotain muuta?
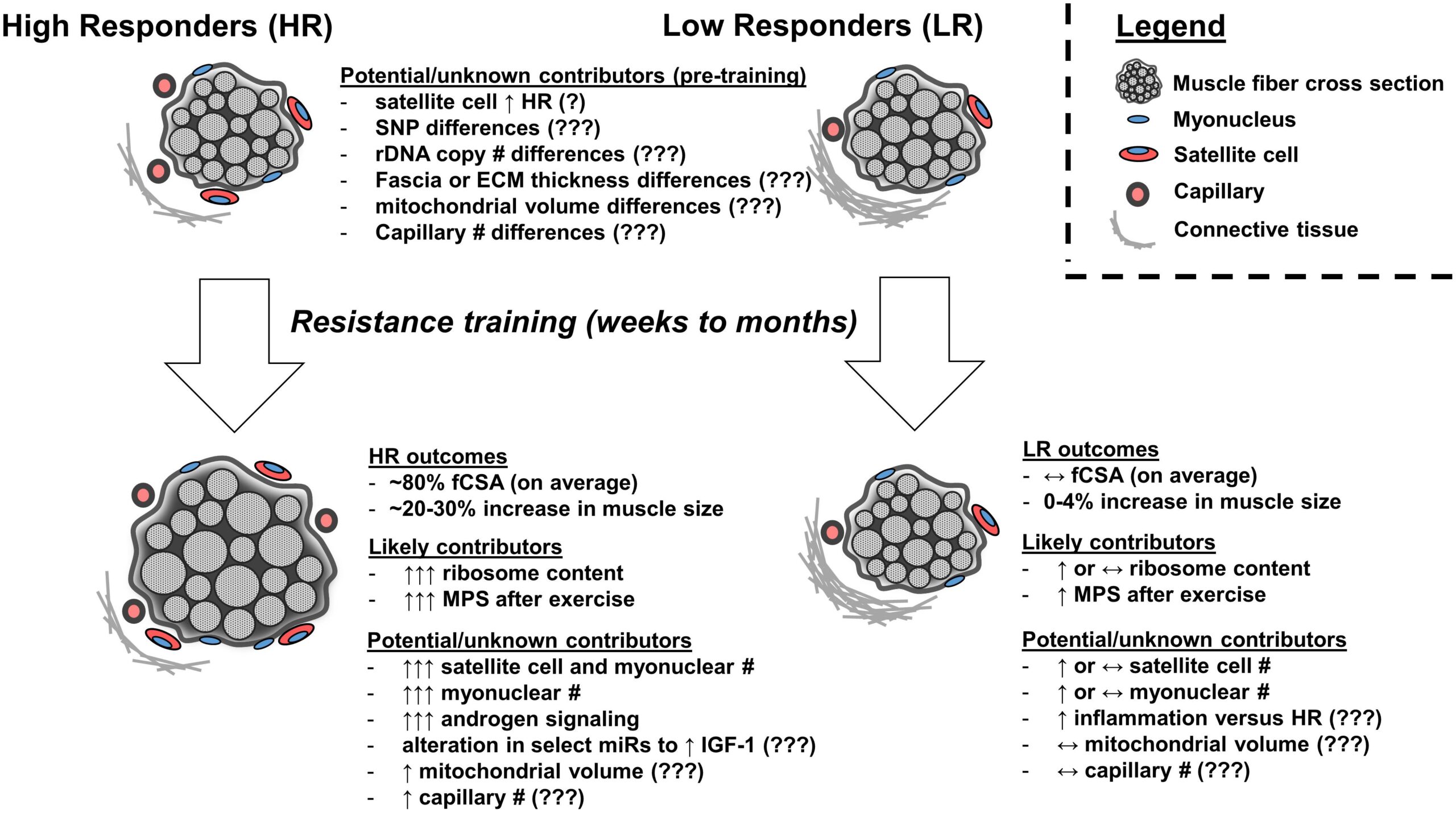
Kuva 1: Nopeasti kehittyvillä yksilöillä näyttäisi olevan suurempi ribosomipitoisuus ja lihasproteiinisynteesi harjoitusten jälkeen verrattuna heikosti kehittyviin yksilöihin. Lisäksi hypoteettisesti hyvin kehittyvillä on enemmän satelliittisoluja, tumia, androgeenisignalointia, mitokondrioiden volyymia ja hiusverisuonistoa. Kuvan lähde Roberts et al., 2018.
Elämäntavat tärkeämpiä kuin geenit?
Genetiikka on eittämättä tärkeässä roolissa yksilön kehityksessä. Esimerkiksi ACTN3 geenin on näytetty olevan yhteydessä eroihin lihasvoiman ja tehontuoton kehittymisessä harjoittelussa (Jones et al., 2016; Eynon et al., 2013). Toisaaltaidenttisillä kaksosilla huomattiin, että elintavoilla on valtava merkitys. Identtiset kaksoset ovat perimältään ja genotyypiltään täysin samanlaiset. Perimän ollessa identtinen, vain olosuhteet vaikuttavat kehitykseen ja näin kaksostutkimukset toimivat esimerkkinä olosuhteiden vaikutuksesta yksilön kehitykseen. 52 vuotiaista kaksosista toinen oli harjoitellut kestävyystyyppisesti yli 30 vuotta ja toinen kaksosista ei ollut liikkunut juuri lainkaan. Harjoitelleella kaksosella oli pienempi kehonmassa, rasvamassa, leposyke, verenpaine, kolesteroli, plasman triglyseridi- ja glukoosipitoisuus. Lisäksi hänellä oli korkeampi anaerobinen ja aerobinen kestävyyskyky. Toisaalta hänellä oli vähemmän lihasta ja voimaa verrattuna ei-harjoitelleeseen kaksoseen. Hänellä oli enemmän hitaita lihassoluja ja vähemmän nopeita lihassoluja sekä korkeampi AMPK proteiinin esiintyvyys (kestävyyssignalointiin liitetty väylä). Yhteensä kestävyysharjoitelleella kaksosella oli 55% enemmän hitaita lihassoluja ja jopa 12,4 ml/kg/min korkeampi VO2max ja 8,6% matalampi rasvaprosentti.
Tutkimuksessa korostettiin elintapojen ja harjoittelun voimakasta vaikutusta koko kehoon. Geenit määrittävät tietyn taustan harjoittelulle, mutta riittävällä harjoittelulla ja elintavoilla voidaan vaikuttaa erittäin tehokkaasti haluttuihin adaptaatioihin. Myös muissa kaksoistutkimuksissa on huomattu (Marsh et al., 2020), etteivät geenit välttämättä ole ihan niin tärkeässä roolissa harjoitusadaptaatiossa kuin aikaisemmin on arvioitu eri poikittaistutkimuksissa.
Toisaalta Davidsenin ja kumppanien tutkimus osoitti, että mikroiRNA-muutokset alhaisen vasteen ihmisillä voivat estää kasvua ja uudistumista edistävien geenien “aktivoitumisen”. Heidän mukaansa lihaskasvu liitty mikroRNA pitoisuuksien muutoksiin ja heidän analyysinsa viittaa siihen, että mikroRNA:lla voi olla rooli voimaharjoittelun aiheuttamissa muutoksissa ja siinä kehittyykö ihminen vai ei. Näyttäisikin siltä, että niin geeneillä kuin olosuhteilla on merkitystä.
Kuva 2: Intohimon lisäksi myös geenit ja olosuhteet vaikuttavat harjoitusadaptaatioiden määrään.
Mitä tehdä jos kehitystä ei tule - lisää harjoituskuormaa?
Harjoituskuormaa, eli volyymia, on pidetty tärkeimpänä tekijänä morfologisia muutoksia varten (Figueiredo et al., 2018). Mattocks et al. (2017) huomasivat, että koehenkilöt, jotka suorittivat hyvin korkean intensiteetin voimaharjoituksia saavuttivat saman voimatasojen kasvamisen kuin ne, jotka harjoittelivat suuremmalla volyymilla. Kuitenkin vain korkeamman volyymin harjoitteluryhmä saavutti lihasmassan kasvua.
Molekyylitason näkökulmasta suurempi harjoitusvolyymi vaikuttaa myofibrillaarisen proteiinisynteesin anabolisen signalointiin positiivisesti. Suurempi volyymi on myös tärkeä p70S6k- ja S6-fosforylaation kannalta voimaharjoittelun jälkeen (Burd ym., 2010a, b; Terzis ym., 2010). p70S6k- ja S6-fosforylaatio säätelevät muun muassa solujen kasvua vaikuttamalla proteiinisynteesin komponentteihin.
Kuinka paljon volyymia on tarpeeksi?
Meta-analyysi ikääntyvien ihmisten lihasmassaharjoittelusta osoitti, että merkittävämmät muutokset liittyivät erityisesti harjoitusohjelmiin, jossa tehtiin enemmän sarjoja koko harjoitussession aikana (Peterson ym., 2011). Useissa tutkimuksissa on huomattu useamman sarjan ylivoimaisuus verrattuna yhteen sarjaan sekä nuorilla että vanhemmilla yksilöillä (Kramer, 1997; Radaelli ym., 2014a,b). Myös Kriegerin (2010) tekemässä meta-analyysissä huomattiin, että usean sarjan suorittaminen on todella tehokkaampaa kuin yhden sarjan suorittaminen. Toisaalta samassa analyysissä huomattiiin, että neljän tai kuuden sarjan suorittaminen ei ole parempaa kuin kahden tai kolmen sarjan suorittaminen. Eli harjoitusvolyymin lisäyksellä voisi olla teoreettinen maksimi yksilön kohdalla. Lisäksi vaikuttaa siltä, ettei supistustapa ole yhtä tärkeä kuin volyymi, jos halutaan saavuttaa molekyylitason muutoksia harjoitussession jälkeen (Garma ym., 2007).
Tietääksemme vain yksi tutkimus on yrittänyt vastata suoraan siihen, voidaanko harjoitusvolyymilla vaikuttaa yksilöllisiin eroihin. Nunes ym. (2021b) huomasivat, että heikosti kehittyvät ihmiset eivät kehittyneet, vaikka harjoitusvolyymia nostettiin. Toisaalta tämä oli retrospektiivinen data-analyysi, eikä suora interventiotutkimus. Scarpelli et al. (2020) näyttivät myös, että ei-optimi harjoitusvolyymi haittaa lihaskasvua nuorilla harjoitelleilla yksilöillä. Toisaalta Montero & Lunby (2017) huomasivat mielenkintoisessa tutkimuksessaan, että kaikki heikosti harjoitteluun vastanneet yksilöt muuttuivat positiivisesti kehittyneiksi yksilöiksi kun harjoittelua lisättiin runsaasti kahdelle seuraavalla harjoitusviikolle kuuden harjoitusviikon jälkeen. Toisaalta tutkimuksessa suoritettiin aerobista harjoittelua, eikä voimaharjoittelua.
Hammarströmin ja kumppanien (2019) erittäin mielenkiintoisessa tutkimuksessa huomattiin korkeamman volyymin kasvattavan enemmän lihasta ja voimaa verrattuna pienempään harjoituskuormaan. Harjoittelu toteutettiin niin, että vastakkainen jalka teki vähemmän volyymia ja toinen jalka enemmän. Harjoituskuorma oli hyvin pieni, mutta ero kolminkertainen (1 sarja vs 3 sarjaa). 13 osallistujaa hyötyi selvästi enemmän suuremmasta volyymista lihasmassan kohdalla ja 16 osallistujaa voiman kohdalla. Ainoastaan kolme osallistujaa hyötyi pienemmästä harjoituskuormituksesta. Lisäksi yksilötasolla korkeampiharjoitusvolyymi oli yhteydessä lisääntyneeseen ribosomien biogeneesiin eli uudimuodostukseen. Myös muut ovat todenneet saman asian (Figueiredo et al. 2015; Stec et al. 2016; Mobley et al. 2018).
Oletko ravi- vai työhevonen?
Lihassolujakauma ei selitä yksin yksilöllisiä eroja voimaharjoittelussa, mutta erittäin mielenkiintoisesti Van Vossel et al. (2023) huomasivat, että saadakseen saman lihasmassan kasvun aikaan enemmän hitaita lihassoluja omaavien pitää tehdä enemmän työtä. Tämä tukee ajatusta siitä, että hidassolukkoisten yksilöiden voi olla kannatavaa tehdä suurempaa volyymiikuormaa. Kannattaa siis rohkeasti kokeilla suurempaa volyymia, jos et kehity. Jotkut yksilöt ovat kuin dieselkoneita, jotka vaativat enemmän kuormaa lähteäkseen kunnolla käyntiin.
Mistä sitten tietää mikä on sopiva harjoituskuorma? Kannattaa aloittaa määrittämällä vähimmäisannos, jolla pystyy kehittymään. Tämän pohjalta voi lähteä etsimään optimaalista harjoituskuormitusannosta (Fisher ym., 2017).
Jos kehitystä ei tule, niin vaihda harjoitusmuotoa
Harjoitusmuodon vaihtamisella voi myös tulla kehitystä. Esimerkiksi tässä mielenkiintoisessa tutkimuksessa 16 rugbypelaajaa tekivät neljä erilaista reeniä, jolla pyrittiin määrittämään testosteronivaste jokaisen eri harjoituksen kohdalla. Tarkoituksena oli mitata, mikä harjoitusmuoto tuotti suurimman ja pienimmän testosteronivasteen. 8 pelaajaa suoritti kolmen viikon harjoitusblokin, jossa tehtiin sitä harjoitusmuotoa, joka tuotti maksimaalisen testosteronivasteen ja toinen porukka suoritti saman pituisen harjoituspätkän, mutta tehden sitä harjoitusmuotoa, joka tuotti pienimmän testosteronivasteen.
Harjoitusliikkeet olivat samat, eli penkki, jalkaprässi, alatalja ja kyyky, mutta sarjat ja toistot vaihtelivat:
Harjoitusmuoto 1: 4 x 10 x 70% 2min palautus.
Harjoitusmuoto 2: 3 × 5 x 85% 3min palautus.
Harjoitusmuoto 3: 5 × 15 x 55% 1min palautus.
Harjoitusmuoto 4: 3 × 5 x 40% 3min palautus.
Kaikki pelaajat kehittyivät merkittävästi penkissä ja jalkaprässissä kun he tekivät harjoitusmuotoa, joka aiheutti suurimman testosteronivasteen. Toisaalta pienimmän hormonaalisen vasteen aiheuttanut harjoitusmuoto ei aiheuttanut mitään muutoksia jopa 75% urheilijoista ja joillakin yhden toiston maksimi jopa pieneni.
Mielenkiintoisesti molemmilla henkilöillä, joilla oli suurin testosteronivaste 3 × 5 x 40% protokollalle, esiintyi 4 × 10 x 70% protokolla vähäisimpänä vasteenaan sekä ennen että jälkeen harjoitusjakson. Samalla tavalla seitsemästä yksilöstä, jotka vastasivat optimaalisesti 4 × 10 x 70% protokollalle, viidelle (71%) 5 × 15 x 55% protokolla tuotti vähiten testosteronivastetta. Tutkimus osoittaa hyvin, että joillekin erilaiset harjoitusmuodot näyttäisivät toimivat paremmin kuin toisille. Onkin tärkeä etsiä juuri yksilölle sopiva harjoitusmuoto.
Henkinen jaksaminen ja vahvuus voivat edesauttaa harjoittelua
Talsnesin ja kumppanien tutkimuksessa pystyttiin erottelemaan korkeasti ja heikosti harjoitteluun vastaavia kestävyysurheilijoita kuuden kuukauden harjoittelujakson jälkeen. Mielenkiintoisesti tutkimuksessa oli haastateltu valmentajia, että mistä erot voisivat johtua. Valmentajien mukaan korkeampi motivaatio ja vahvemmat valmentaja-urheilija suhteet korkean vasteen ryhmässä vaikuttivat yksilöllisesti optimoitujen harjoitus- ja palautumisrutiinien käyttöön, mikä johti positiivisempaan suorituskyvyn kehitykseen. Tämä johti myös siihen, että korkean vasteen ryhmän urheilijat suorittivat suurempia harjoitusmääriä (viikoittainen kuormitus: 3825 ± 1013 vs. 3228 ± 748 ja kuormitus/volyymi-suhde: 4,9 ± 0,6 vs. 4,2 ± 0,5; molemmat P ≤ 0,05) ja heillä oli vähemmän loukkaantumisia/sairastumisia verrattuna heikosti kehittyviin (5 ± 3 vs. 10 ± 5 päivää; P = 0,07).
Yhteenveto
Yksilöiden välillä on runsaasti eroja kehittymisessä
Ei kannata selittää kehittymättömyyttään geeneillä. Geenit asettavat tietyt reunaehdot, mutta kehitys on mahdollista kaikille.
Joillekin yksilöille sopii suurempi harjoituskuorma kuin toisille. Yksilöllisen vasteprofiilin etsiminen on tärkeää.
Erilaisia harjoitusmuotoja kokeilemalla voi myös saada kehitystä aikaan.
Lähteet
Montero,D., and Lundby, C. (2017). Refuting the myth of non-response to exercise training: ’non-responders’ do respond to higher dose of training. J. Physiol. 595, 3377–3387. doi: 10.1113/JP273480
Nunes, J. P., Pina, F. L., Ribeiro, A. S., Cunha, P. M., Kassiano, W., Costa, B. D., et al. (2021b). Responsiveness to muscle mass gain following 12 and 24 weeks of resistance training in older women. Aging Clin. Exp. Res. 33, 1071–1078. doi: 10.1007/s40520-020-01587-z
Scarpelli, M. C., Nóbrega, S. R., Santanielo, N., Alvarez, I. F., Otoboni, G. B., Ugrinowitsch, C., et al. (2020). Muscle hypertrophy response is affected by previous resistance training volume in trained individuals. J. Strength Cond. Res. 27, 1–5. doi: 10.1519/JSC.0000000000003558
Fisher, J. P., Steele, J., Gentil, P., Giessing, J., andWestcott,W. L. (2017). A minimal dose approach to resistance training for the older adult; the prophylactic for aging. Exp. Gerontol., 99, 80–86. doi: 10.1016/j.exger.2017.09.01
Kramer, J. B. (1997). Effects of single vs multiple sets of weight training: impact of volume, intensity, and variation. J. Strength Cond. Res. 11, 143–147. doi: 10.1519/00124278-199708000-00002
Radaelli, R., Botton, C. E., Wilhelm, E. N., Bottaro, M., Brown, L. E., Lacerda, F., et al. (2014a). Time course of low- and high-volume strength training on neuromuscular adaptations and muscle quality in older women. Age 36, 881–892. doi: 10.1007/s11357-013-9611-2
Radaelli, R.,Wilhelm, E. N., Botton, C. E., Rech, A., Bottaro,M., Brown, L. E., et al. (2014b). Effects of single vs. multiple-set short-term strength training in elderly women. Age 36:9720. doi: 10.1007/s11357-014-9720-6
Krieger, J. W. (2010). Single vs. multiple sets of resistance exercise for muscle hypertrophy: a meta-analysis. J. Strength. Cond. Res. 24, 1150–1159. doi: 10.1519/JSC.0b013e3181d4d436
Jones, N., Kiely, J., Suraci, B., Collins, D. J., de Lorenzo, D., Pickering, C., … & Grimaldi, K. A. (2016). A genetic-based algorithm for personalized resistance training. Biology of sport, 33(2), 117.
Eynon, N., Hanson, E. D., Lucia, A., & Houweling, P. J. (2013). Genes for elite power and sprint performance: ACTN3 leads the way. Sports Medicine, 43(9), 803-817.
Rhea, M. R., Alvar, B. A., Burkett, L. N., & Ball, S. D. (2003). A meta-analysis to determine the dose response for strength development. Medicine and science in sports and exercise, 35(3), 456-464.
Phillips, S. M., & Van Loon, L. J. (2011). Dietary protein for athletes: from requirements to optimum adaptation. Journal of sports sciences, 29(sup1), S29-S38.
Burd, N. A., Holwerda, A.M., Selby, K. C.,West, D.W., Staples, A.W., Cain, N. E., et al. (2010a). Resistance exercise volume affects myofibrillar protein synthesis and anabolic signalling molecule phosphorylation in young men. J. Physiol. 588, 3119–3130. doi: 10.1113/jphysiol.2010.192856
Terzis, G., Spengos, K., Mascher, H., Georgiadis, G., Manta, P., and Blomstrand, E. (2010). The degree of p70 S6k and S6 phosphorylation in human skeletal muscle in response to resistance exercise depends on the training volume. Eur. J. Appl. Physiol. 110, 835–843. doi: 10.1007/s00421-010-1527-2
Garma, T., Kobayashi, C., Haddad, F., Adams, G. R., Bodell, P. W., and Baldwin, K. M. (2007). Similar acute molecular responses to equivalent volumes of isometric, lengthening, or shortening mode resistance exercise. J. Appl. Physiol. 102, 135–143. doi: 10.1152/japplphysiol.00776.2006
Peterson,M. D., Sen, A., and Gordon, P.M. (2011). Influence of resistance exercise on lean body mass in aging adults: a meta-analysis. Med. Sci. Sports Exerc. 43, 249–258. doi: 10.1249/MSS.0b013e3181eb626
Hammarström, D., Øfsteng, S., Koll, L., Hanestadhaugen, M., Hollan, I., Apró, W., Whist, J.E., Blomstrand, E., Rønnestad, B.R. and Ellefsen, S. (2020), Benefits of higher resistance-training volume are related to ribosome biogenesis. J Physiol, 598: 543-565. https://doi.org/10.1113/JP278455
Figueiredo VC, Caldow MK, Massie V, Markworth JF, Cameron-Smith D & Blazevich AJ (2015). Ribosome biogenesis adaptation in resistance training-induced human skeletal muscle hypertrophy. Am J Physiol Endocrinol Metab 309, E72– E83.
Mobley CB, Haun CT, Roberson PA, Mumford PW, Kephart WC, Romero MA, Osburn SC, Vann CG, Young KC, Beck DT, Martin JS, Lockwood CM & Roberts MD (2018). Biomarkers associated with low, moderate, and high vastus lateralis muscle hypertrophy following 12 weeks of resistance training. PLoS One 13, e0195203.
Stec MJ, Kelly NA, Many GM, Windham ST, Tuggle SC & Bamman MM (2016). Ribosome biogenesis may augment resistance training-induced myofiber hypertrophy and is required for myotube growth in vitro. Am J Physiol Endocrinol Metab 310, E652– E661.
Beaven, C Martyn1; Cook, Christian J2; Gill, Nicholas D1. Significant Strength Gains Observed in Rugby Players after Specific Resistance Exercise Protocols Based on Individual Salivary Testosterone Responses. Journal of Strength and Conditioning Research 22(2):p 419-425, March 2008. | DOI: 10.1519/JSC.0b013e31816357d4
Talsnes RK, van den Tillaar R, Cai X, Sandbakk Ø. Comparison of High- vs. Low-Responders Following a 6-Month XC Ski-Specific Training Period: A Multidisciplinary Approach. Front Sports Act Living. 2020 Sep 8;2:114. doi: 10.3389/fspor.2020.00114. PMID: 33345103; PMCID: PMC7739740.
Bathgate KE, Bagley JR, Jo E, Talmadge RJ, Tobias IS, Brown LE, Coburn JW, Arevalo JA, Segal NL, Galpin AJ. Muscle health and performance in monozygotic twins with 30 years of discordant exercise habits. Eur J Appl Physiol. 2018 Oct;118(10):2097-2110. doi: 10.1007/s00421-018-3943-7. Epub 2018 Jul 14. PMID: 30006671.
Marsh CE, Thomas HJ, Naylor LH, Scurrah KJ, Green DJ. Fitness and strength responses to distinct exercise modes in twins: Studies of Twin Responses to Understand Exercise as a THerapy (STRUETH) study. J Physiol. 2020 Sep;598(18):3845-3858. doi: 10.1113/JP280048. Epub 2020 Jul 7. PMID: 32567679.
Davidsen PK, Gallagher IJ, Hartman JW, Tarnopolsky MA, Dela F, Helge JW, Timmons JA, Phillips SM. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J Appl Physiol (1985). 2011 Feb;110(2):309-17. doi: 10.1152/japplphysiol.00901.2010. Epub 2010 Oct 28. PMID: 21030674.
Van Vossel, K., Hardeel, J., de Casteele, F.V., Van der Stede, T., Weyns, A., Boone, J., Blemker, S., Lievens, E. and Derave, W. (2023), Can muscle typology explain the inter-individual variability in resistance training adaptations?. J Physiol. Accepted Author Manuscript. https://doi.org/10.1113/JP284442
Figueiredo VC, de Salles BF, Trajano GS. Volume for Muscle Hypertrophy and Health Outcomes: The Most Effective Variable in Resistance Training. Sports Med. 2018 Mar;48(3):499-505. doi: 10.1007/s40279-017-0793-0. PMID: 29022275.
Mattocks KT, Buckner SL, Jessee MB, Dankel SJ, Mouser JG, Loenneke JP. Practicing the Test Produces Strength Equivalent to Higher Volume Training. Med Sci Sports Exerc. 2017 Sep;49(9):1945-1954. doi: 10.1249/MSS.0000000000001300. PMID: 28463902.
Isometrisellä lihastyötavalla viitataan lihastyöhön, missä lihasjännekompleksin pituudessa ei tapahdu muutosta. Konsentrisessa lihastyötavassa lihas lyhenee supistuessaan ja eksentrisessä lihastyötavassa lihas pitenee lihassolujen supistuessa. Esimerkkinä kyykystä ylös ponnistaminen on konsentrista lihastyötä suurimmalla osalla jalkojen lihaksia ja lihasten pituus lyhenee, kun taas eksentrinen toiminta pidentää lihaspituutta. Tästä esimerkkinä, kun mennään alaspäin kyykyssä. Isometristä harjoittelua voidaan käytännössä tehdä monella eri tavalla, mutta tässä jutussa keskitytään pelkästään työskentelyyn liikkumatonta esinettä vasten.
Isometrinen harjoittelu on erityisen mielenkiintoinen aihealue urheilijoille, koska isometristä harjoittelua voi hyödyntää positiivisten hermolihasjärjestelmän adaptaatioiden saavuttamiseksi ilman liiallista väsymystä, mitä perinteinen keskiraskas voimaharjoittelu aiheuttaa.
VIDEO 1: Yleisin tapa toteuttaa käytännössä isometristä voimaharjoittelua liikkumatonta esinettä vasten on käyttää tankoa ja suojarautoja tai vastaavia tukirakenteita.
Minkälaisia adaptaatiota isometrinen harjoittelu aiheuttaa
Voiko pelkällä isometrisellä harjoittelulla kasvattaa lihasta?
Kyllä voi! Isometrinen harjoittelu 42–100 päivän ajan on johtanut 5,4–23% lihaksen poikkipinta-alan kasvuun ja jopa 91,7% nousuun maksimivoimassa (28-37). Pidempikestoinen interventio näyttäisi vaikuttavan huomattavasti lihaksen kokoon. Mitä pidempi interventio oli, sitä enemmän lihas kasvoi. Hypertrofiaan vaikutti myös harjoittelun intensiteetti, voluumi, supistuksen kesto ja lihaksen pituus.
Erityisesti pitkillä lihaspituuksilla tehty isometrinen harjoittelu parantaa ylivoimaisesti enemmän lihaksen kokoa verrattuna lyhyillä lihaspituuksilla tehtyyn isometriseen harjoitteluun, vaikka volyymi olisi tasattu näiden ryhmien välillä (1, 2 & 3). Tulokset ovat lähes samansuuntaisia, kun verrataan isometristä harjoittelua dynaamiseen harjoitteluun. Myös normaalissa dynaamisessa voimaharjoittelussa näyttäisi laaja liikerata olevan huomattavasti hyödyllisempi hypertrofian kannalta verrattuna vajaisiin liikeratoihin (4, 5 & 6). Yksi syy tähän voi olla, että pitkällä lihaspituudella tehdyt supistukset näyttäisivät tuottavan huomattavasti enemmän lihasvaurioita verrattuna lyhyellä lihaspituudella tehtäviin harjoitteisiin (7). Tämä johtuu siitä, että nivelen vipuvarsi kasvaa pitkillä lihaspituuksilla ja näin lisää mekaanista jännitystä lihaksessa verrattuna lyhyempään vipuvarteen. Suurempi mekaaninen jännitys aiheuttaa enemmän lihasvaurioita. Lisäksi pitkät lihaspituudet kuluttavat enemmän happea, vaativat enemmän verenkierrolta töitä ja kokonaisuudessaan lisäävät metaboliittien kerääntymistä enemmän kuin lyhyet lihaspituudet (8). Metaboliset tekijät ovat tutkitusti myös yhteydessä lihaskasvuun (9). Eli jos tavoitteena on spesifi lihaskasvu isometrisessä harjoittelussa, niin pitkät lihaspituudet ovat ehdottomasti paras valinta.
Volyymillä on selvästi väliä myös isometrisessä harjoittelussa, kun tavoitteena on lihaskasvu. Meyers (10) vertaili matala volyymista harjoittelua (3 x 6 sekuntia maksimaalisella intensiteetillä) korkea volyymiseen harjoitteluun (20 x 6 sekuntia maksimaalisella intensiteetillä) hauislihaksella. Kuuden viikon jälkeen enemmän volyymia tehnyt ryhmä oli saavuttanut selvästi isomman muutoksen hauislihaksen ympärysmitassa verrattuna matalavolyymiseen ryhmään. Myös Balshaw ja kumppanit (11) totesivat, että suurempi määrä volyymia (40 x 3 sekuntia 75% isometrisestä maksimista) tuotti enemmän lihaskasvua etureiteen 12-viikon aikana verrattuna pienempään harjoituskuormaan (40 x 1 sekuntia 80% isometrisestä maksimista).
Mielenkiintoista on myös, että Schott ja kumppanit (12) löysivät, että pidempikestoinen harjoittelu (4 x 30 sekuntia) tuotti enemmän hypertrofiaa verrattuna lyhyempikestoiseen harjoitteluun (4 x 10 x 3 sekuntia), vaikka liikesuoritteiden kokonaiskesto oli lopulta sekunnilleen yhtä pitkä. 14-viikon harjoittelun jälkeen etureiden vastus lateralis lihas kasvoi jopa 11,1% enemmän, kun lyhyempikestoisia supistuksia tehneellä ryhmällä ei löydetty ollenkaan merkitsevää muutosta etureiden kasvusta! Tämä voi johtua siitä, että pitkään ylläpidetyt supistukset estävät verenkierron ja vähentävät hapen saturaatiota alueella, stimuloiden näin hypertrofiaa monien paikallisten ja systeemisten mekanismien kautta.
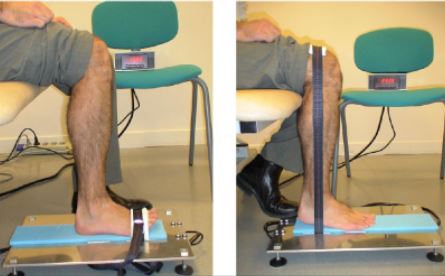
KUVA 1: Isometristä harjoittelua voidaan tutkia nilkan plantaari- tai dorsifleksiota tarkkailemalla tämän näköisellä koeasetelmalla.
Isometrinen harjoittelu muokkaa myös lihaksen arkkitehtuuria
Hypertrofiaa haettaessa lihastyötavalla ei ole hirveästi merkitystä, sillä niin dynaamisella, eksentrisellä ja isometrisellä harjoittelulla voidaan saada lihaskasvua aikaiseksi, mutta jos tavoitteena on saada muutoksia aikaan lihaksen arkkitehtuuriin, on lihatyötavalla todellakin merkitystä.
Laadukkaita tutkimuksia aiheesta ei ole paljoa, joten päätelmien tekeminen on haastavaa, mutta Noorkoiv ja kumppanit (3) huomasivat, että pidemmällä lihaspituudella tehty isometrinen harjoittelu (polvikulma 38.1 ± 3.7°) kasvatti vastus lateraliksen lihasfasciculuksen (lihassolukimppu, jota ympäröi lihaskalvo) pituutta keskiosassa lihasta merkitsevästi. Mielenkiintoisesti lyhyemmällä lihaspituudella tehty harjoittelu kasvatti taas distaalisessa päässä olevan lihasfasciculuksen pituutta. Ainoastaan yksi toinen tutkimus (1) on raportoinut vastus lateraliksen lihasfasciculuksen pituuden lisääntymistä ja myös pennaatiokulman muutoksesta pitkällä lihaspituudella tehdyn isometrisen harjoittelun jälkeen.
Isometrinen voimaharjoittelu näyttäisi aiheuttavan muutoksia lihaksen arkkitehtuuriin ja erityisesti lisäävän lihasfasciculuksen pituutta ja kenties jopa aiheuttaa muutoksia pennaatiokulmaan. Tällä on erityisesti väliä, jos tavoitteena on tehdä urheilijoista nopeampia, sillä esimerkiksi sprinttereillä on pidemmät lihasfasciculukset jaloissa verrattuna kestävyysurheilijoihin (38) ja 100 metrin juoksusuoritus on yhdistetty lihasfasciculuksien pituuksiin (39).
Isometrisen harjoittelun vaikutukset jänteisiin
Jänteen tarkoitus on siirtää voimia luun ja lihaksen välillä mahdollistaen nivelen liike. Ennen ajateltiin jänteiden olevan muuttumattomia, mutta onneksi nykyään tiedetään jo, että jänteet kykenevät adaptoitumaan stimulukseen merkitsevästi ja voivat käydä todella isoja arkkitehtuurisia muutoksia läpi pitkäaikaisen kuormituksen johdosta.
Esimerkiksi kun vertaillaan eri lajien urheilijoita akillesjännerepeämän kokemiin ihmisiin, on huomattu, että esimerkiksi lentopalloilijoilla on huomattavasti suurempi akillesjänne (119 ± 5.9) verrattuna akillesjännerepeämän kokemiin ihmisiin (101 ± 5.4). Mielenkiintoista oli, että kajakkiurheilijoilla oli lähes samankokoinen akillesjänne kuin repeämän kokemilla ihmisillä (101 ± 5.6) (13). Kajakkiurheilijat eivät juuri käytä akillesjänteitään lajissaan, joten harjoittelulla näyttäisi olevan suuri vaikutus jänteen rakenteisiin.
Jänteen adaptaatiot ovat erittäin tärkeitä ja haluttuja adaptaatioita nopeuslajin urheilijoille, sillä jänne toimii nopeassa liikkeessä liikuttajana jousen tavoin. Intensiteetti on ehdottomasti tärkein muuttuja jänteen adaptaatioissa. Kova intensiteettinen isometrinen plantaarifleksion harjoittelu (noin 90 % isometrisestä maksimista) lisäsi akillesjänteen poikkipinta-alaa ja jäykkyyttä 14-viikon harjoitteluohjelman aikana jopa parhaimmillaan 36 % (14 & 15). Samaa ei huomattu matalaintensiteettisellä harjoittelulla (55 % isometrisestä maksimista). Myös muut ovat raportoineet samankaltaisia runsaita muutoksia jänteen jäykkyydessä (vaihteluväli 17,5 % - 61,6 %) isometrisen voimaharjoittelun seurauksena intensiteetin vaihdellessa 70–100 % välillä isometrisestä maksimivoimasta (16, 17 & 18). Näyttäisi siltä, että 70 % voisi olla minimi-intensiteetti, joka vaaditaan haluttujen jänneadaptaatioiden saavuttamiseksi.
Räjähtävä isometrinen voimaharjoittelu taas lisäsi jänteen aponeuroosin elastisuutta, mutta vähensi jänteen poikkipinta-alaa (-2,8 %) (19). Isometrisen harjoittelun intensiteetillä ja kestolla saavutetaan hyvin erilaisia adaptaatioita. Jänteiden vahvistamisessa tulee suosia pidempiä ja intensiteetti korkealla tehtyjä supistuksia, kun taas kisakaudella voi tehdä terävämpiä elastisuutta lisääviä erittäin lyhyitä supistuksia. Lisäksi pidempi lihaspituus näyttäisi kehittävän jänteen jäykkyyttä enemmän kuin harjoittelu lyhyellä lihaspituudella samalla tavalla kuin lihaskasvussa (2).
KUVA 2: Bruce Leekin käytti isometristä harjoittelua kehittämään omaan suorituskykyään.
Isometrisen voimaharjoittelun vaikutukset hermostoon
Hermoston adaptaatiot ovat kokonaisuudessaan hyvin harjoitteluspesifejä. Esimerkiksi Balshaw ja kumppanit (11) vertailivat 12 viikon aikana maksimaalista voimaharjoittelua (1 sekunnin rauhallinen nousu 75% isometrisestä maksimista ja siellä 3s pito) räjähtävään voimaharjoitteluun (mahdollisimman nopeasti >90% isometriseen maksimiin ja siellä 1s pito). Isometrinen maksimivoima kehittyi eniten maksimivoimaharjoittelulla, mutta räjähtävä voimaharjoittelu lisäsi EMG aktiivisuutta ihan liikkeen alussa (0–100 ms aikana) enemmän verrattuna maksimivoimaharjoitteluun. Nämä adaptaatiot olivat hermostoperäisiä ja olivat harjoitteluspesifejä, kun maksimivoimaharjoittelu kehitti maksimivoimaa ja räjähtävä voima kehitti nopeaa voimantuottokykyä. Myös ballistinen isometrinen harjoittelu on johtanut samankaltaisiin tuloksiin ja EMG amplitudin paranemiseen ensimmäisen 0-150 ms aikana verrattuna maksimivoimaharjoitteluun (11, 23 & 24).
Isometrisellä voimaharjoittelulla voidaan vaikuttaa lihaksen jännitys-pituussuhteeseen, eli siihen, millä lihaksenpituudella tai nivelen kulmalla tuotetaan isoin mahdollinen voima. Tämä on erityisen tärkeä urheilussa, jossa halutaan maksimoida suurin mahdollinen tuotettu voima halutussa asennossa. Myös parasta voimantuottokulmaa voidaan muokata isometrisellä harjoittelulla. Esimerkiksi Alegre ja kumppanit (25) raportoivat, että pidemmällä lihaspituudella harjoittelu kahdeksan viikon ajan johti 11 asteen muutokseen kohti pidempiä lihaspituuksia, kun taas lyhyemmillä kulmilla harjoittelu johti 5,3 astetta optimaalista kulmaa toiseen suuntaan. Myös Bogdanis ja kumppanit (26) huomasivat noin 10 % tiputuksen optimaalisessa kulmassa lyhyillä liikeradoilla harjoitellessa.
Pidempikestoinen supistus näyttäisi olevan joissakin tapauksissa tehokkaampi tapa parantaa voimaa ja myös dynaamista urheilun suorituskykyä (hyppäämistä ja juoksemista) verrattuna nopeaan isometriseen voimantuottotapaan (40). Pidemmässä supistuksessa tehtiin kolmen sekunnin ajan työtä ja räjähtävässä nopeassa isometrisessä voimantuottotavassa tehtiin yhden sekunnin verran töitä. Tuloksia on tulkittava hieman varovasti, sillä pidempää supistusta tehnyt ryhmä teki yhteensä 15 sekunnin verran työtä sarjassa, kun lyhyempää pätkää tehnyt ryhmä teki vain 10 sekunnin verran työtä. Kuuden viikon aikana ja 12 harjoituskerran vuoksi erot kertaantuvat ja tehty kokonaistyö oli huomattavasti isompi kolmen sekunnin ryhmässä verrattuna yhden sekunnin ryhmään. Tämä varmasti osaltaan selittää tuloksia.
Kolmen sekunnin ryhmä paransi esikevennettyä hyppyä 12,1 % ja yhden sekunnin ryhmä 10,8 %. Erot kasvaneista voimatasoissakin voivat selittää nämä muutokset. Mielenkiintoisesti pidempikestoinen isometrinen voimaharjoittelu aiheutti 1,4 % parannuksen 30 metrin juoksuaikaan. Tässäkin tapauksessa enemmän harjoitellut ryhmä paransi huomattavasti enemmän nopeuttaan, kun vähemmän harjoitellut ryhmä. Voisiko kasvaneet voimatasot, ei niinkään nopeus, selittää erot. Normaalilla kovalla kyykyllä ja plyometrisellä harjoittelulla on saatu 1,2 % parannus 30 metrin juoksuaikaan (43), joka on aika lähellä tämän tutkimuksen saamia tuloksia.
Toisaalta tässäkin tutkimuksessa huomattiin, että kyky tuottaa voimaa nopeammin parani yhden sekunnin ryhmällä enemmän kuin kolmen sekunnin ryhmällä, kun taas pidempikestoisessa supistuksessa maksimivoima kehittyi enemmän. Myös muut ovat raportoineet samankaltaisia tuloksia (41 & 42).
Yhteenveto
Isometristä harjoittelua voi hyödyntää positiivisten hermolihasjärjestelmän adaptaatioiden saavuttamiseksi ilman liiallista väsymystä. Tämä on erityisen tärkeää erityisesti urheilijoilla kilpailukauden aikana. Lisäksi jos tiettyä voimantuottokulmaa tai lajin vaatimia kulmia pitää harjoitella, niin isometrinen harjoittelu on erittäin tehokas työkalu niihin.
Isometrinen harjoitteluun pätee samat lainalaisuudet kuin muuhunkin harjoitteluun. Hypertrofiaa saavuttaaksesi tulee harjoittelua tehdä 70-75% intensiteetillä maksimaalisesta supistuksesta noin 3-30s ajan toistossa ja sarjamäärän ollessa > 80 – 150s per yksi harjoituskerta. Maksimivoimaa saavuttaaksesi isometristä harjoittelua tulee tehdä 80-100% maksimaalisesta supistuksesta 1-5s ajan ja kokonaiskeston ollessa 30-90s. Voimantuottonopeutta parantaakseen tulee suorituksessa pyrkiä tuottamaan mahdollisimman nopeasti mahdollisimman paljon voimaa. Sarjan keston tulee olla lyhyt. Kuvassa 3 on koottu tämänhetkiseen tutkimusnäyttöön perustuen ohjeistus isometriseen voimaharjoitteluun.
KUVA 3: Tämänhetkisestä tukimusnäytöstä koostettu taulukko miten isometristä harjoittelua tulisi tehdä, jos haluaa saavuttaa tietyn adaptaation.
Tiivistys
Lihastakin voin kasvattaa pelkällä isometrisellä harjoittelulla. Volyymi ja lihaspituus ovat tärkeimmät muuttujat, kun tavoitteena on lihaskasvu.
Isometrisessä harjoittelussa intensiteetti on päämuuttuja voiman kohdalla. Hypertrofian kohdalla volyymi.
Ballistisella protokola on ylivoimainen räjähtävän voiman kehittymiseen. Ensimmäiselle 50 ja 100 ms voi parantaa voimantuottoa huomattavasti. Jos tämä on tavoite, niin harjoitteet tulisi tehdä mahdollisimman nopeasti ja mahdollisimman voimakkaasti.
Lähteet:
Alegre LM, Ferri-Morales A, Rodriguez-Casares R, Aguado X. Effects of isometric training on the knee extensor moment–angle relationship and vastus lateralis muscle architecture. Eur J Appl Physiol. 2014;114(11):2437-2446.
Kubo K, Ohgo K, Takeishi R, et al. Effects of isometric trainingmat different knee angles on the muscle–tendon complex in vivo. Scand J Med Sci Sports. 2006;16(3):159-167.
Noorkoiv M, Nosaka K, Blazevich AJ. Neuromuscular adaptations associated with knee joint angle-specific force change. Med Sci Sports Exerc. 2014;46(8):1525-1537.
Guex K, Degache F, Morisod C, Sailly M, Millet GP. Hamstring architectural and functional adaptations following long vs. short muscle length eccentric training. Front Physiol. 2016;7(340):1-9.
Barak Y, Ayalon M, Dvir Z. Transferability of strength gains from limited to full range of motion. Med Sci Sports Exerc. 2004;36(8):1413-1420.
Massey CD, Vincent J, Maneval M, Moore M, Johnson JT. An analysis of full range of motion vs. partial range of motion training in the development of strength in untrained men. J Strength Cond Res. 2004;18(3):518-521.
Allen TJ, Jones T, Tsay A, Morgan DL, Proske U. Muscle damage produced by isometric contractions in human elbow flexors. J Appl Physiol. 2018;124(2):388-399.
de Ruiter CJ, de Boer MD, Spanjaard M, de Haan A. Knee angle-dependent oxygen consumption during isometric contractions of the knee extensors determined with near-infrared spectroscopy. J Appl Physiol. 2005;99:579-586.
Dankel SJ, Mattocks KT, Jessee MB, Buckner SL, Mouser JG, Loenneke JP. Do metabolites that are produced during resistance exercise enhance muscle hypertrophy? Eur J Appl Physiol. 2017;117(11):2125-2135.
Meyers CR. Effects of two isometric routines on strength, size, and endurance in exercised and nonexercised arms. Res Q Exerc Sport. 1967;38(3):430-440
Balshaw TG, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specific functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol. 2016;120(11):1364-1373.
Schott J, McCully K, Rutherford OM. The role of metabolites in strength training: short versus long isometric contractions. Eur J Appl Physiol Occup Physiol. 1995;71(4):337-341.
Kongsgaard M, Aagaard P, Kjaer M, Magnusson SP. Structural Achilles tendon properties in athletes subjected to different exercise modes and in Achilles tendon rupture patients. J Appl Physiol (1985). 2005 Nov;99(5):1965-71. doi: 10.1152/japplphysiol.00384.2005. Epub 2005 Aug 4. PMID: 16081623.
Arampatzis A, Karamanidis K, Albracht K. Adaptational responses of the human Achilles tendon by modulation of the applied cyclic strain magnitude. J Exp Biol. 2007;210:2743-2753.
Arampatzis A, Peper A, Bierbaum S, Albracht K. Plasticity of human Achilles tendon mechanical and morphological properties in response to cyclic strain. J Biomech. 2010;43(16):3073-3079.
Burgess KE, Connik MJ, Graham-Smith P, Pearson SJ. Plyometric vs isometric training influences on tendon propertied and muscle output. J Strength Cond Res. 2007;21(3):986-989.
Kubo K, Kanehisa H, Fukunaga T. Effects of different duration isometric contractions on tendon elasticity in human quadriceps muscles. J Physiol. 2001;536(2):649-655.
Kubo K, Ishigaki T, Ikebukuro T. Effects of plyometric and isometric training on muscle and tendon stiffness in vivo. Physiol Rep. 2017;5(e13374):1-13
Massey G, Balshaw T, Maden-Wilkinson T, Tillin N, Folland J. Tendinous tissue adaptation to explosive- vs. sustained-contraction strength training. Front Physiol. 2018;9(1170):1–17.
Bandy WD, Hanten WP. Changes in torque and electromyographic activity of the quadriceps femoris muscles following isometric training. Phys Ther. 1993;73(7):455-465.
Barak Y, Ayalon M, Dvir Z. Transferability of strength gains from limited to full range of motion. Med Sci Sports Exerc. 2004;36(8):1413-1420.
Massey CD, Vincent J, Maneval M, Moore M, Johnson JT. An analysis of full range of motion vs. partial range of motion training in the development of strength in untrained men. J Strength Cond Res. 2004;18(3):518-521.
Tillin NA, Folland JP. Maximal and explosive strength training elicit distinct neuromuscular adaptations, specific to the training stimulus. Eur J Appl Physiol. 2014;114(2):365-374.
Maffiuletti NA, Martin A. Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Med Sci Sports Exerc. 2001;33(7):1220-1227
Alegre LM, Ferri-Morales A, Rodriguez-Casares R, Aguado X. Effects of isometric training on the knee extensor moment– angle relationship and vastus lateralis muscle architecture. Eur J Appl Physiol. 2014;114(11):2437-2446.
Bogdanis GC, Tsoukos A, Methenitis SK, Selima E, Veligekas P, Terzis G. Effects of low volume isometric leg press complex training at two knee angles on force-angle relationship and rate of force development. Eur J Sport Sci. 2018;1-9. https://doi.org /10.1080/17461391.2018.1510989. [Epub ahead of print].
Behm DG, Sale DG. Intended rather than actual movement velocity determines velocity-specific training response. J Appl Physiol. 1993;74(1):359-368.
Balshaw T, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specific functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol (1985) 2016; 120: 1364–1373
Davies J, Parker DF, Rutherford OM, Jones DA. Changes in strengh and cross sectional area of the elbow flexors as a result of isometric strength training. Eur J Appl Physiol 1988; 57: 667–670
Garfinkel S, Cafarelli E. Relative changes in maximal force, EMG, and muscle cross-sectional area after isometric training. Med Sci Sports Exerc 1992; 24: 1220–1227
Ikai M, Fukunaga T. A study on training effect on strength per unit corss-sectional area of muscle by means of ultrasonic measurement. Eur J Appl Physiol 1970; 28: 173–180
Jones DA, Rutherford OM. Human muscle strength training: The effects of three different regimes and the nature of the resultant changes. J Physiol 1987; 391: 1–11
Kanehisa H, Nagareda H, Kawakami Y, Akima H, Masani K, Kouzaki M, Fukunaga T. Effects of equivolume isometric training programs comprising medium or high resistance on muscle size and strength. Eur J Appl Physiol 2002; 87: 112–119
Kubo K, Ohgo K, Takeshi R, Yoshinaga K, Tsunoda N, Kanehisa H, Fukunaga T. Effects of isometric training at different knee angles on the muscle-tendon complex in vivo. Scand J Med Sci Sports 2006; 16: 159–167
Noorkoiv M, Nosaka K, Blazevich AJ. Neuromuscular adaptations associated with knee joint angle-specific force change. Med Sci Sports Exerc 2014; 46: 1525–1537
Noorkoiv M, Nosaka K, Blazevich AJ. Effects of isometric quadriceps strength training at different muscle lengths on dynamic torque production. J Sports Sci 2015; 33: 1952–1961
Schott J, McCully K, Rutherford OM. The role of metabolites in strength training II. Short vs. long isometric contractions. Eur J Appl Physiol 1995; 71: 337–341
Abe, Takashi, Kumagai, Kenya, Brechue, William F. Fascicle length of leg muscles is greater in sprinters than distance runners, Medicine & Science in Sports & Exercise: June 2000; 32(6): 1125-1129.
Kumagai K, Abe T, Brechue WF, Ryushi T, Takano S, Mizuno M. Sprint performance is related to muscle fascicle length in male 100-m sprinters. J Appl Physiol (1985). 2000 Mar;88(3):811-6. doi: 10.1152/jappl.2000.88.3.811. PMID: 10710372.
Lum, D., Barbosa, T.M., Joseph, R. et al. Effects of Two Isometric Strength Training Methods on Jump and Sprint Performances: A Randomized Controlled Trial. J. of SCI. INSPORTANDEXERCISE3, 115–124 (2021). https://doi.org/10.1007/s42978-020-00095-w
Balshaw T, Massey GJ, Maden-Wilkinson TM, Tillin NA, Folland JP. Training-specifc functional, neural, and hypertrophic adaptations to explosive- vs. sustained-contraction strength training. J Appl Physiol. 2016;120(11):1364–73
Tillin NA, Folland JP. Maximal and explosive strength training elicit distinct neuromuscular adaptations, specifc to the training stimulus. Eur J Appl Physiol. 2014;114(2):365–74.
Ronnestad BR, Kvamme NH, Sunde A, Raastad T. Short-term efects of strength and plyometric training on sprint and jump performance in professional soccer players. J Strength Cond Res. 2008;22(3):773–80
Behm DG, Sale DG (1993) Intended rather than actual movement velocity determines velocity-specific training response. Journal of Applied Physiology 74(1):359–68.
Maffiuletti NA, Martin A (2001) Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Medicine and Science in Sports and Exercise 33(7):1220–27.
Olsen PD, Hopkins WG (2003) The effect of attempted ballistic training on the force and speed of movements. Journal of Strength and Conditioning Research 17(2):291–98.
Viitasalo JT, Komi PV (1981) Effects of fatigue on isometric force- and relaxation-time characteristics in human muscle. Acta Physiologica Scandavica 111(1):87–95.
Oranchuk DJ, Storey AG, Nelson AR, Cronin JB. Isometric training and long-term adaptations: Effects of muscle length, intensity, and intent: A systematic review. Scand J Med Sci Sports. 2019 Apr;29(4):484-503. doi: 10.1111/sms.13375. Epub 2019 Jan 13. PMID: 30580468.
Käy lukemassa vieraskirjoitus Lihastohtorin blogista miten lihasten kokoa ja voimaa voi ylläpitää sekä kehittää ilman pääsyä kuntosalille. Viime viikon harjoittelutauko kirjoituksen jatkoksi julkaistu käytännön osuus on nyt ulkona ja tarjoaa paljon käytännön vinkkejä kuvien ja videoiden muodossa.
Artikkelin yhteenveto:
Tällä hetkellä ei kannata keskittyä siihen mitä kaikkea ei pysty tekemään, vaan sen sijaan mitä pystyy tekemään.
Nyt tämä tilanne pakottaakin monet monipuolistamaan harjoittelua ja antaa mahdollisuuden oppia paljon uusia liikkeitä ja tapoja harjoitella.
Ilman kuntosalin laitteita ja välineitä voi harjoitella ja vähintäänkin ylläpitää saavutettuja voima- ja lihasmassatasoja.
Kehittävää tai vähintäänkin ylläpitävää harjoittelua voi tehdä omalla kehonpainolla yksin tai parin kanssa ja tarvittaessa erilaisia yksinkertaisia välineitä käyttäen.
Vastustuskyvyn kannalta olisi hyvä, jos harjoittelu ei olisi intensiteetiltään ja määrältään kehoa toistuvasti äärirajoille vievää nykyisessä epidemiassa.
Käy lukemassa vieraskirjoitus Lihastohtorin blogista harjoittelemattomuusjakson vaikutuksesta lihaksen kokoon ja voimaan.
Artikkelin yhteenveto:
Alle neljän viikon harjoittelemattomuusjakso ei vaikuta voimatasoihin juuri ollenkaan varsinkin kokeneemmilla ja vanhemmilla treenaajilla. Tämän jälkeen voima alkaa vähitellen vähenemään.
Lihakset alkavat hiljalleen surkastua noin kolmen viikon treenitauon jälkeen.
Treenitauko saattaa liioitella lihasten pienentymistä. Älä säikähdä sitä, sillä tämä johtuu osittain lihasten glykogeenivarastojen pienentymisestä ja viimeisen treenin aiheuttaman pienen lihasturvotuksen katoamisesta, ei ”oikean” lihasmassan häviämisestä. Tämä vaikutus on väliaikainen, ja jo pienikin harjoittelu palauttaa glykogeenitasot lihaksessa.
Pidemmän harjoittelemattomuusjakson jälkeen jälleen aloitettu harjoittelu palauttaa menetetyn lihasmassan ja voiman takaisin hyvin nopeasti ”lihasmuistin” ansiosta.
Lihaskasvua käsittelevä artikkelisarja lähenee loppua ja on viimeisen osan vuoro. Tässä artikkelissa käsitellään miten eri yksilöt adaptoituvat voimaharjoitteluun ja miten harjoittelu kannattaisi ohjelmoida, jotta kehitys olisi kaikkein optimaalisinta.
Genetiikan vaikutus lihaskasvuun
Geenit vaikuttavat suuresti henkilön kehittymiseen. Teoreettisesti voidaan ajatella maksimaalisen lihaskasvun rajojen olevan olemassa. Nämä määrittyvät genotyypin, eli geneettisen informaation, ja näiden geenien ilmentymisen yksilössä mukaan. Geenit asettavat teoreettisesti meille tietyt reunaehdot, miten reagoimme erilaisiin tilanteisiin. Geenien takia jotkut yksilöt kehittyvät nopeammin ja jotkut yksilöt hitaammin. Kuiinka paljon vaihtelua voi sitten eri henkilöiden välillä olla?
KUVA 1: Tarkistuslista, jos epäilet salimarkun tai salijonnan käyttävän kehon ulkopuolisia hormoneita.
Yksilölliset vasteet harjoitteluun vaihtelevat hurjasti ihmisillä. Tämä tarkoittaa sitä, että jos kaksi samaa ihmistä tekee täysin saman ohjelman niin tuskin koskaan he kehittyvät täsmälleen saman verran. Esimerkiksi samanlaisella harjoittelulla lihaksen kasvu voi vaihdella jopa -11 % - + 30 % välillä ja voima taas - 8 % - + 60 % välillä (Ahtiainen et al., 2016). Ei-respondoivia ihmisiä, eli niitä, jotka eivät reagoi mitenkään tutkimusten voimaharjoitteluun on keskimäärin 25% koehenkilöistä (Bamman et al., 2007). Toisaalta nykyään ajatellaan, että erilainen harjoittelu, ravinto, lepo tai joku muu muuttuja olisi voinut vaikuttaa yksilöllisiin vasteisiin. Eli tälläiset ei-respondoivat koehenkilöt johtuvat pääosin huonosta tutkimusasetelmasta, elämäntilanteesta tai muusta tekijästä, mikä estää kehityksen. Jotkut ei-respondoivista ihmisistä tarvitsevat vain enemmän ja pidempää harjoittelua verrattuna toisiin.
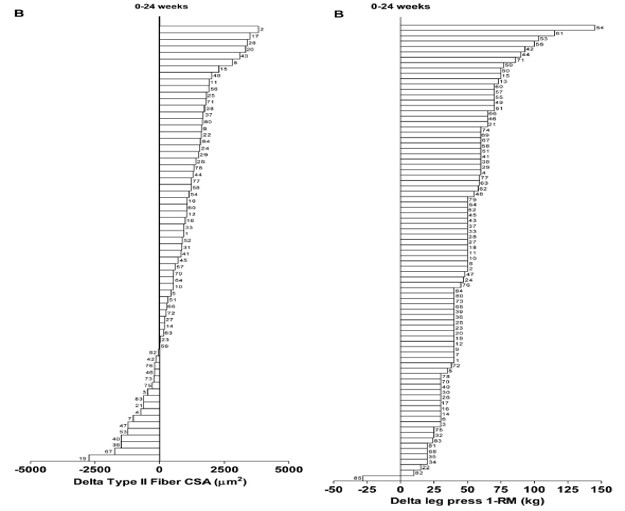
KUVA 2: Oikeassa kaaviossa nähdään, että lähes kaikki koehenkilöt saivat erään tutkimuksen aikana lisää voimaa, mutta vasemmalla huomataan, että joidenkin koehenkilöiden lihaskoko jopa pieneni tutkimuksen aikana. (Churward-Venne et al., 2015).
Mikä näitä eroja sitten selittää? Pescatello ja kummanit (2013) pystyivät tunnistamaan 17 geeniä, joiden uskotaan selittävän osan henkilöiden välisistä eroista. Näistä tärkeimpänä hypertrofian kannalta pidetään tällä hetkellä BMP2-geeniä (Devaney et al., 2009). Ja myös MGF:ää pidetään erittäin tärkeänä (Bamman et al., 2007).
Lisäksi satelliittisolut lisääntyvät ja luovuttavat tumia tehokkaammin hyvin respondoituvilla ihmisillä kuin verrataan ei-respondoituviin ihmisiin (Petrella et al., 2008). Myös lihaksen morfologia vaikuttaa suuresti lihaskasvuun. Mitä enemmän lihassoluja henkilöllä on, niin sen helpommin lihas kasvaa. Lihassolujen määrä on aika vakio ja hyperplasiaa (lihassolujen lisääntymistä) ei tapahdu enää 24 viikon jälkeen syntymästä. Lihassolujen määrä onkin pitkälti geenien määrittämä.
Jos vertaillaan mieskehonrakentajia ja tavallisia kadun tallaajia niin kehonrakentajilla on enemmän lihassoluja hauiksissa ja nämä solut olivat myös isompia verrattuna tavallisiin tallaajiin (MacDougall et al., 1984). Joten niillä, jotka päätyivät kehonrakentajiksi oli jo enemmän lihassoluja kuin muilla normaaleilla ihmisillä. Tämä on yksi syy, miksi kehonrakentajat reagoivat paremmin voimaharjoitteluun.
Nopeiden lihassolujen määrä kasvatettavassa lihaksessa on tärkeä tekijä sen suhteen miten voimaharjoittelu vaikuttaa lihakseen. Tyypin II - lihassolut kasvavat noin 50% enemmän verrattua tyypin I -lihassoluihin. Toisaalta tässäkin suhteessa on hurjasti henkilöiden välistä vaihtelua (Kosek et al., 2006). Tutkimuksia, jotka selittävät fysiologisia mekanismeja yksilöllisessä voimaharjoittelussa on saatavilla erittäin niukasti, mutta erot johtuvat yleisesti perimästä, geenien ilmenemisessä, soluviestinnässä, hormonireseptoreiden toiminnassa ja satelliittisoluissa. Nostan vielä esille ravitsemuksen, jota harvoin tutkimuksissa kontrolloidaan ja sillä on valtava merkitys lihaskasvun mahdollistamisessa.
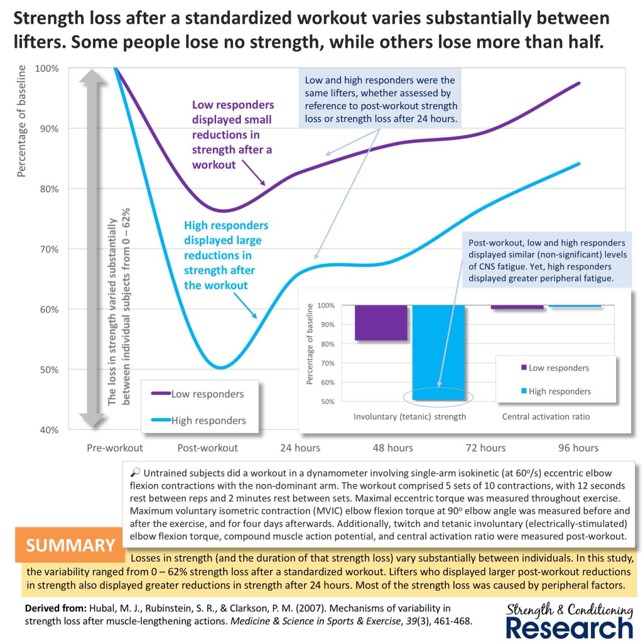
KUVA 3: Näyttäisi myös siltä, että hyvin voimaharjoitteluun reagoivat ihmiset saavat itsestään irti paljon harjoituksen aikana. Heillä harjoituksen jälkeen voima laskee enemmän kuin huonosti respondoituvilla ihmisillä. Tämä tarkoittaa sitä, että he ovat saaneet enemmän väsymystä aikaiseksi harjoituksen aikana.
Genetiikalla on siis kieltämättä iso rooli henkilön lihaskasvussa. Voit kiittää vanhempiasi näistä lahjoista, mutta muuten asiaan ei kannata kiinnittää sen enempää huomiota. Jokainen voi ja pystyy kehittymään, vaikka tutkimuksissa usein löydetään ei-respontoivia henkilöitä harjoitteluun niin tähän saattaa olla syynä vääränlainen tutkimuksenasettelu, harjoittelu tai ohjelmointi. Yksilöllinen harjoittelu ja ohjelma mahdollistavat jokaisen kehittymisen. Heikosti respondoivien pitää joskus reenata jopa paljon vähemmän ja harvemmin kehittyäkseen. Tai kevyemmillä kuormilla. Myös harjoitustausta vaikuttaa suuresti henkilön kykyyn adaptoitua. Aloittelevilla ihmisillä lihaskasvu voi olla jopa 28% ja korkeimmillaan 58% 16 viikossa (Stewart & Rittweger, 2006), kun taas kehonrakentajat kehittyivät viidessä kuukaudessa “vain” 3-7% (Alway et al., 1992). Ohjelmointi ja ohjelman muuttujien manipulointi korostuu erityisesti, kun harjoitustaustaa kertyy enemmän. Kevennetyt jaksot voivat auttaa taas respontoimaan paremmin stimulukseen (Ogasawara et al., 2013).
Ei-respondereilla ei siis ole mielenvika pääsään vaan se on ihan oikea asia. Heidän tulee erityisesti keskittyä ravintoon ja oikeanlaiseen harjoitteluun. Huonosti respondoivat saattavat olla vain hitaammin respondoivia ja tarvitsevat enemmän harjoitusvolyymia kehittyäkseen. Nimi on vähän harhaanjohtava sillä jokainen kehittyy oikeanlaisella harjoittelulla. Saatat vain olla heikommin ja hitaammin respondoituva henkilö verrattuna salikaveriisi. Lisäksi vielä useissa tutkimuksissa kehitys on 12 viikossa esim. non respondereilla “vain” 5%. Tämä tulos luokitellaan tutkimuksissa hitaasti reaspondoivaksi, mutta jos kehitys jatkuisi koko vuoden verran samanlaisena se vastaisi alle vuodessa esimerkiksi 100kg penkissä siirtymistä 121,6 kiloon. Kuinka moni olisi tyytyväinen tuohon kehitykseen vuodessa? Minä ainakin olisin. Joten nimi on hiukan harhaanjohtava ja yleisesti ongelmat johtuvat tutkimusasetelmasta, sekä heterogeenisestä tutkimusjoukosta.
Neuvostoliitossa
periodisaation (ja ylipäätään liikuntatieteen) tutkimus- ja kehitystyö oli
tuolloin huomattavan laajaa ja systemaattista. Tämän tutkimustyön tuloksena
syntyi klassinen periodisaatiomalli. Muissakin maissa toki käytettiin ja
kehitettiin samantyylistä periodisaatiota, tosin pienemmässä mittakaavassa.
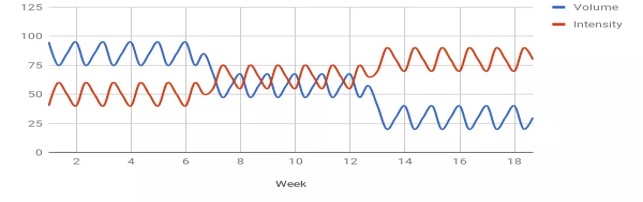
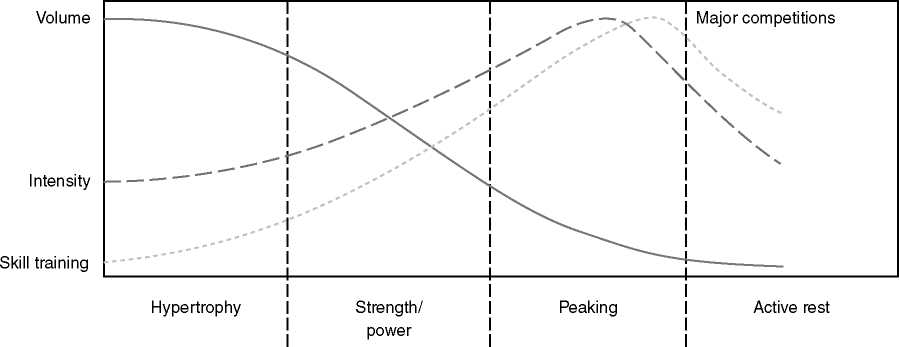
KUVA 4: Klassinen periodisaatiomalli, missä harjoitusvolyymi on aluksi korkealla ja vähenee pääkilpailun lähestyessä. Samalla harjoittelun intensiteetti nousee.
Periodisaatio
on erittäin vähän tutkittu aihealue. Johtuen luultavasti siitä, että
periodisaatio ei ole mikään selkeästi käsitettävä kokonaisuus. Erillaisia
malleja voidaan esittää, mutta näidenkin sisällä voi olla loputtamattomasti
erillaisia muutujia, joita voi siirrellä ja muutella. Periodisaatio perustuu
Selyen yleiseeen adaptaatio syndrooma - teoriaan (GAS - general adaptation
syndrome). Se perustuu kehon reaktioihin, jotka ovat hälytys, vastustus ja
uupuminen. (Selye, 1950). Teoriaa on nykypäivinä kritisoitu
yksinkertaistamisesta ja siitä, että se ei huomioi esimerkiksi psykologisia,
teknisiä ja ravinnollisia seikkoja. Selyen teorian mukaan harjoittelua
pidetään stressitekijänä, johon voidaan soveltaa GAS - teoriaa. Harjoittelun
tulee olla progressiivisesti haastavaa, jotta tasannevaihetta ei tule
harjoittelussa ja toisaalta harjoittelu ei saa olla ylikuluttavaa.
Periodisaatiomalleista kolmea on tutkittu lihashypertrofian kannalta: perinteinen lineaarinen malli, ei-lineaarinen ja käänteinen periodisaatio.
Perinteisessä
lineaarisessa mallissa on vähintään makrosykli, mesosykli ja mikrosykli
(esimerkiksi vuosi, peruskuntokausi 8 vko, yksi harjoitusviikko). Malli etenee
korkeasta voluumista ja matalasta intensiteetistä kohti korkeaan
intensiteettiin ja matalaan voluumiin. Esimerkiksi rakenteellinen
jakso/hypertrofinen jakso, jossa voluumi nousee ja intensiteetti pysyy 60-75%
välillä, seuraavaksi voi olla perusvoimakausi intensiteetin ollessa 80-90%
välillä ja voluumin vähetessä. Viimeinen mesosykli olisi intensiteetiltään
suurin 90-100%, mutta voluumiltaan pienin.
Lihaskasvun osalta aihetta on tutkittu jonkin verran, mutta itse pidän tutkimuksia metodologisilta osuuksilta ontuvilta, joten johtopäätöksiä ei tule vetää vielä näiden tutkimusten pohjalta. Stone ja kumppanit (1981) tutkivat periodisaatioidun ohjelman ja ei-periodisoidun ohjelman eroja, ja huomasivat hypertrofian (mitattu veden alaispunnituksella) eroavan tilastollisesti merkitsevästi ei-periodisoidusta ohjelmoinnista. Baker:in tutkimusporukka taa (1994) ei löytänyt eroja periodisatioidun ja ei-periodisaatiouidun välille (ihopoimumittaus). Kun taas Monteiro ja kumppanit (2009) löysivät eron (ihopoimu, ei tilastollisesti merkittävä) periodisoidun harjoittelun hyväksi.
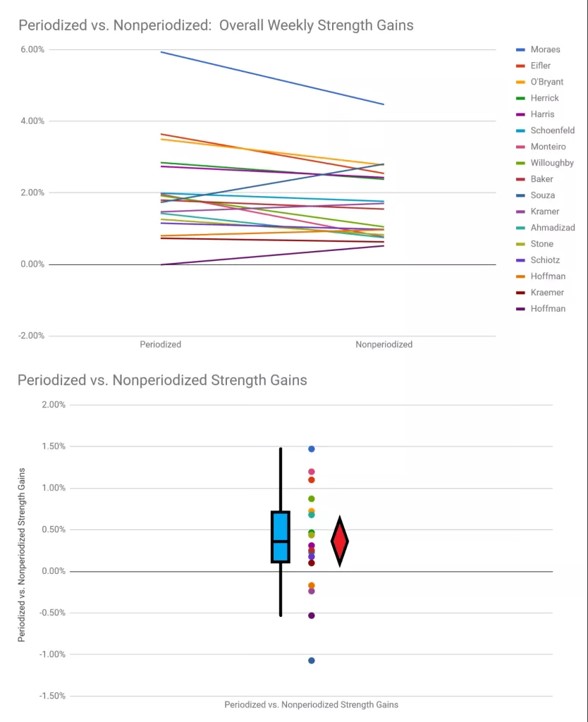
Voiman osalta periodisoitu harjoittelu tuottaa kiistatta parempia tuloksia kuin ei periodisoitu harjoittelu (Ahmadizad et al., 2014;, Monteiro et al., 2009,Willoughby, 1993). Voisi ajatellakin, että koska voima nousee paremmin periodisoidulla harjoittelulla, johtaisi se mekaanisen kuormituksen johdosta myös suurempiin hypertrofisiin adaptaatioihin ajan kanssa, mutta tätä ei ole vielä todistettu.
KUVA 5: Periodisoitu harjoittelu tuottaa keskimäärin 21,78 – 23,62% voiman nousun, kun taas ei periodisoitu harjoittelu johtaa vain 18,90 – 19,10% keskimääräiseen voiman nousuun. Viikkotasolla samat luvut 1,96 % - 2,05 % verrattuna ei periodisoituun 1,59 – 1,70%. (Nuckols, 2018.)
Ei-lineaarinen periodisaatio tai aaltoileva periodisaatio on nykyään yksi suosituimmista suuntauksista. Sitä on käytetty jo kauan, mutta Poliquin (1988) oli ensimmäinen, joka esitteli sen tiedeyhteisössä. Hänen mukaansa lineaarisen ohjelmoinnin ongelmat ovat liian suuri progressiivinen painon lisäys liian lyhyessä ajassa ja alussa saavutettujen hypertrofisten adaptaatioiden menetys muissa mesosykleissä, koska voluumi laskee liikaa.
Aaltoilevassa periodisaatiossa vaihdellaan voluumia ja intensiteettiä kuinkas muutenkaan kuin aaltoilevaan tyyliin. Mallia on kehitetty eteenpäin ja tällä hetkellä suosituin malli taitaa olla päivittäin vaihtuva periodisaatio (daily undulating periodization - DUP). Tässä mallissa on viikon sisällä maksimivoima, perusvoima ja hypertrofinen harjoitus. Aihetta on tutkittu suhteellisen laajasti. Ja vain yksi tutkimus sai tilastollisesti merkitsevän eron lineaarisen ja ei-lineaarisen mallin välille. Tässä tutkimuksessa ei-lineaarinen malli oli selvästi parempi lisäämään kyynärpään koukistajien ja ojentajien tiheyttä (Simao et al., 2012). Muuten muissa tutkimuksissa ei ole saatu eroja mallien välille lihaskasvun osalta, mikä osoittaa kummankin mallin olevan yhtä toimiva lihaskasvun osalta (Baker et al., 1994; Davitt et al., 2014; , Harries et al., 2015; Monteiro et al., 2009, Prestes et al., 2009, Simao et al., 2012). Toisaalta tutkimusten kesto on ollut korkeimmillaan vain 12 viikkoa ja kun puhutaan periodisaatiosta yleisesti urheilijoiden osalta puhutaan vuosista.
Itse suosin valmennettavillani enemmän hybridimallia, joka on versio aaltoilevasta mallista. Tässä mallissa harjoitusohjelma on laadittu blokkeihin ja näiden blokkien sisällä tapahtuu aaltoilua. Blokit toisaalta etenevät lineaarisesti. Viikon sisällä siinä tehdään maksimivoimasarjoja ja perusvoima sarjoja samassa harjoituksessa järkevästi ohjelmoiden. Pitkillä perusvoimakausilla saattaa hävitä liikaa maksimivoimaa ja pitkillä maksimivoima kausilla saattaa hävitä liikaa lihasta tai perusvoimaa, joten järkevä hybridiohjelmointi poistaa nämä ongelmat.
KUVA 6: Aaltoileva periodisaatio, missä volyymi ja intensiteetti aaltoilevat mahdollistaen intensiteetin ylläpidon peruskuntokaudella ja volyymin ylläpidon kisakaudella.
Käänteistä periodisaatiota on ehdotettu lihaskasvuun tähtääville tärkeimmäksi ohjelmaksi. Lineaarinen malli tiputtaa jatkuvasti voluumia kohti kauden päätähtäintä ja kun tiedetään voluumin olevan elintärkeä hypertrofian kannalta voisi ajatella lineaarisen mallin olevan huono lihaskasvun kannalta. Käänteisessä mallissa voluuumi on korkeimmillaan kauden päätähtäimen kohdalla. Aihe on vielä tutkimuskentällä tuore ja vain tietääkseni Prestes ja kumppanit (2009) ovat sitä tutkineet huonoin tuloksin käänteisen mallin puolesta. Lineaarinen malli oli huomattavasti parempi kuin käänteinen malli. Toisaalta mittauksissa käytettiin ihopoimupihtejä, jotka ovat metodisesti yhtä tyhjän kanssa.
Tiettyä suunnitelmallisuutta harjoitteluun tarvitaan, koska muuten harjoittelusta puuttuu variaatio, progressiivisuus ja tavoitteet. Lihaskasvun osalta yksi tärkeimmistä periodisaation tuomista asioista on kevennetyt jaksot. Keho ottaa paremmin vastaan harjoittelua kuin välillä tiputtaa voluumia ja antaa kropan palautua. Lisäksi on tärkeä kerätä palautetta periodisaation toimivuudesta ja suunnitelman muokkaaminen sen mukaan auttavat kehittämään ohjelmointia.
Kiitos lukijalle, jos jaksoit lukea tänne saakka! Alla lähdeluettelo kaikista artikkelisarjan osista. Aiheeseen liittyvä lukeminen ei ihan heti lopu kesken. Jätä palautetta tai kommentteja ja tsemppiä treeneihin!
Lähteet:
Ahtiainen, J., Pakarinen, A., Kraemer, W. &
Häkkinen, K. (2003). Acute
Hormonal and Neuromuscular Responses and Recovery to Forced vs. Maximum
Repetitions Multiple Resistance Exercises. International Journal of Sports
Medicine, 24(6), 410–418. doi: 10.1055/s-2003-41171
Ahtiainen, J. P.,
Pakarinen, A., Alen, M., Kraemer, W. J., & Häkkinen, K. (2005). Short vs. Long Rest Period Between the Sets in
Hypertrophic Resistance Training: Influence on Muscle Strength, Size, and
Hormonal Adaptations in Trained Men. The Journal of Strength and Conditioning
Research, 19(3), 572. doi: 10.1519/15604.1
Ahtiainen JP, Walker S, Peltonen H, Holviala J,
Sillanpää E, Karavirta L, et al. Heterogeneity in resistance
training-induced muscle strength and mass responses in men and women of
different ages. Age (Omaha). 2016;38:1–13.
Ahmadizad, S., Ghorbani, S., Ghasemikaram, M. &
Bahmanzadeh, M. (2014) Effects of short-term nonperiodized, linear periodized
and daily undulating periodized resistance training on plasma adiponectin,
leptin and insulin resistance. Clinical biochemistry, 47: 417-422.
Alegre, L.,
Jimenez, F., Gonzalo-Orden, J., Acero, R. & Aguado, X. (2006). Effects of
dynamic resistance training on fascicle length and isometric strength. Journal
of sports sciences: 24. 501-8. 10.1080/02640410500189322.
Allen, D. G.,
Whitehead, N. P., & Yeung, E. W. (2005). Mechanisms of stretch-induced
muscle damage in normal and dystrophic muscle: role of ionic changes. The
Journal of Physiology, 567(3), 723–735. doi: 10.1113/jphysiol.2005.091694
Alway, S. E.,
Grumbt, W. H., Stray-Gundersen, J., & Gonyea, W. J. (1992). Effects of
resistance training on elbow flexors of highly competitive bodybuilders.
Journal of Applied Physiology, 72(4), 1512–1521. doi:
10.1152/jappl.1992.72.4.1512
Atherton, P. J.,
Babraj, J., Smith, K., Singh, J., Rennie, M. J., & Wackerhage, H. (2005).
Selective activation of AMPK-PGC-1α or PKB-TSC2-mTOR signaling can explain
specific adaptive responses to endurance or resistance training-like electrical
muscle stimulation. The FASEB Journal, 19(7), 786–788. doi:
10.1096/fj.04-2179fje
Baker, D., Wilson, G. &
Carolyn, R. (1994) Periodization: The effect on strength of manipulating volume
and intensity. Journal of strength and conditioning research, 8: 235-242.
Bamman, M. M.,
Petrella, J. K., Kim, J.-S., Mayhew, D. L., & Cross, J. M. (2007). Cluster analysis tests the importance of myogenic gene
expression during myofiber hypertrophy in humans. Journal of Applied
Physiology, 102(6), 2232–2239. doi: 10.1152/japplphysiol.00024.2007
Benziane, B.,
Burton, T. J., Scanlan, B., Galuska, D., Canny, B. J., Chibalin, A. V., …
Stepto, N. K. (2008). Divergent cell signaling after short-term intensified
endurance training in human skeletal muscle. American Journal of
Physiology-Endocrinology and Metabolism, 295(6). doi:
10.1152/ajpendo.90428.2008
Blazevich, A. J.,
Cannavan, D., Coleman, D. R., & Horne, S. (2007). Influence of concentric
and eccentric resistance training on architectural adaptation in human
quadriceps muscles. Journal of Applied Physiology, 103(5), 1565–1575. doi:
10.1152/japplphysiol.00578.2007
Bondesen, B. A.,
Mills, S. T., & Pavlath, G. K. (2006). The COX-2 pathway regulates growth
of atrophied muscle via multiple mechanisms. American Journal of
Physiology-Cell Physiology, 290(6). doi: 10.1152/ajpcell.00518.2005
Bondesen, B. A.,
Mills, S. T., Kegley, K. M., & Pavlath, G. K. (2004). The COX-2 pathway is
essential during early stages of skeletal muscle regeneration. American Journal
of Physiology-Cell Physiology, 287(2). doi: 10.1152/ajpcell.00088.2004
Brentano, M. &
Martins Kruel, L. (2011) A review on strength exercise-induced muscle damage:
applications, adaption mechanisms and limitations. J Sports Med Phys Fitness,
51(1): 1-10.
Burd, N. A.,
Holwerda, A. M., Selby, K. C., West, D. W. D., Staples, A. W., Cain, N. E., …
Phillips, S. M. (2010). Resistance exercise volume affects myofibrillar protein
synthesis and anabolic signalling molecule phosphorylation in young men. The
Journal of Physiology, 588(16), 3119–3130. doi: 10.1113/jphysiol.2010.192856
Burd, N. A.,
Mitchell, C. J., Churchward-Venne, T. A., & Phillips, S. M. (2012). Bigger
weights may not beget bigger muscles: evidence from acute muscle protein
synthetic responses after resistance exercise. Applied Physiology, Nutrition,
and Metabolism, 37(3), 551–554. doi: 10.1139/h2012-022
Choi J., Takahashi H., Itai Y. (1998) The
difference between effects of ‘power-up type’ and ‘bulk-up type’ strength
training exercises: With special reference to muscle cross-sectional area. Jpn J Phys Fitness Sports Med 47(1), 119-129.
Clarkson, P. M.,
Byrnes, W. C., Mccormick, K. M., Turcotte, L. P., & White, J. S. (1985).
Muscle Soreness And Serum Creatine Kinase Activity Following Isometric,
Eccentric And Concentric Exercise. Medicine & Science in Sports &
Exercise, 17(2), 277. doi: 10.1249/00005768-198504000-00424
Coffey, V. G.,
Zhong, Z., Shield, A., Canny, B. J., Chibalin, A. V., Zierath, J. R., &
Hawley, J. A. (2006). Early signaling responses to divergent exercise stimuli
in skeletal muscle from well-trained humans. The FASEB Journal, 20(1), 190–192.
doi: 10.1096/fj.05-4809fje
Cornelison, D.
& Wold, B. (1997) Single-cell analysis of regulatory gene expression in
quiescent and activated mouse skeletal muscle satellite cells. Advances in
developmental biology, 191(2): 270–283.
Conboy, I. M., Conboy, M. J.,
Wagers, A. J., Girma, E. R., Weissman, I. L., & Rando, T. A. (2005).
Rejuvenation of aged progenitor cells by exposure to a young systemic
environment. Nature, 433(7027): 760–764. doi: 10.1038/nature03260
Dangott, B., Schultz, E.,
& Mozdziak, P. E. (2000). Dietary Creatine Monohydrate Supplementation
Increases Satellite Cell Mitotic Activity During Compensatory Hypertrophy.
International Journal of Sports Medicine, 21(1), 13–16. doi:
10.1055/s-2000-8848
Davitt, P. M., Pellegrino, J.
K., Schanzer, J. R., Tjionas, H., & Arent, S. M. (2014). The Effects of a
Combined Resistance Training and Endurance Exercise Program in Inactive College
Female Subjects. Journal of Strength and Conditioning Research, 28(7),
1937–1945. doi: 10.1519/jsc.0000000000000355
Devaney, J. M., Tosi, L. L.,
Fritz, D. T., Gordish-Dressman, H. A., Jiang, S., Orkunoglu-Suer, F. E., …
Rogers, M. B. (2009). Differences in fat and muscle mass associated with a
functional human polymorphism in a post-transcriptionalBMP2gene regulatory
element. Journal of Cellular Biochemistry, 107(6), 1073–1082. doi:
10.1002/jcb.22209
Dhawan, J., & Rando, T. A.
(2005). Stem cells in postnatal myogenesis: molecular mechanisms of satellite
cell quiescence, activation and replenishment. Trends in Cell Biology, 15(12),
666–673. doi: 10.1016/j.tcb.2005.10.007
Egner, I. M., Bruusgaard, J.
C., Eftestøl, E., & Gundersen, K. (2013). A cellular memory mechanism aids
overload hypertrophy in muscle long after an episodic exposure to anabolic
steroids. The Journal of Physiology, 591(24): 6221–6230. doi:
10.1113/jphysiol.2013.264457
Evans, W. J., & Cannon, J.
G. (1991). The Metabolic Effects of Exercise-Induced Muscle Damage. Exercise
and Sport Sciences Reviews, 19(1). doi: 10.1249/00003677-199101000-00003
Finkenzeller, G., Newsome, W.,
Lang, F., & Häussinger, D. (1994). Increase of c-jun mRNA upon hypo-osmotic
cell swelling of rat hepatoma cells. FEBS Letters, 340(3), 163–166. doi:
10.1016/0014-5793(94)80129-0
Folland, J. &
Williams, A. (2007) The adaptations to strength training - morphological and
neurological contributions to increased strength. Sports medicine, 37 (2):
145-168.
Fonseca, R. M.,
Roschel, H., Tricoli, V., Souza, E. O. D., Wilson, J. M., Laurentino, G. C., …
Ugrinowitsch, C. (2014). Changes in Exercises Are More Effective Than in
Loading Schemes to Improve Muscle Strength. Journal of Strength and
Conditioning Research, 28(11), 3085–3092. doi: 10.1519/jsc.0000000000000539
Fry, C. S., Glynn,
E. L., Drummond, M. J., Timmerman, K. L., Fujita, S., Abe, T., … Rasmussen, B.
B. (2010). Blood flow restriction exercise stimulates mTORC1 signaling and
muscle protein synthesis in older men. Journal of Applied Physiology, 108(5),
1199–1209. doi: 10.1152/japplphysiol.01266.2009
Gardiner NE. Athletics of the ancient world. Oxford:
University Press; 1930.
Glass, D. J.
(2010). PI3 Kinase Regulation of Skeletal Muscle Hypertrophy and Atrophy.
Current Topics in Microbiology and Immunology Phosphoinositide 3-Kinase in
Health and Disease, 267–278. doi: 10.1007/82_2010_78
Goodman, C. A.,
Mayhew, D. L., & Hornberger, T. A. (2011). Recent progress toward
understanding the molecular mechanisms that regulate skeletal muscle mass.
Cellular Signalling, 23(12), 1896–1906. doi: 10.1016/j.cellsig.2011.07.013
Gordon, S. E.,
Kraemer, W. J., Vos, N. H., Lynch, J. M., & Knuttgen, H. G. (1994). Effect
of acid-base balance on the growth hormone response to acute high-intensity
cycle exercise. Journal of Applied Physiology, 76(2), 821–829. doi:
10.1152/jappl.1994.76.2.821
Goto, K., Ishii,
N., Kizuka, T. & Takamatsu, K. (2005) The impact of metabolic stress on
hormonal responses and muscular adaptations. Medicine and science in sports and
exercise, 37(6): 955-963.
Guyton. A. C., Hall J. E. 2011. Textbook of Medical
Physiology. 12th edition. Philadelphia, USA : W.B. Saunders Company
Harries, S. K.,
Lubans, D. R., & Callister, R. (2016). Comparison of resistance training
progression models on maximal strength in sub-elite adolescent rugby union
players. Journal of Science and Medicine in Sport, 19(2), 163–169. doi:
10.1016/j.jsams.2015.01.007
Hackett, D. A.,
Johnson, N. A., & Chow, C.-M. (2013). Training Practices and Ergogenic Aids
Used by Male Bodybuilders. Journal of Strength and Conditioning Research,
27(6), 1609–1617. doi: 10.1519/jsc.0b013e318271272a
Handayaningsih,
A.-E., Iguchi, G., Fukuoka, H., Nishizawa, H., Takahashi, M., Yamamoto, M., …
Takahashi, Y. (2011). Reactive Oxygen Species Play an Essential Role in IGF-I
Signaling and IGF-I-Induced Myocyte Hypertrophy in C2C12 Myocytes.
Endocrinology, 152(3), 912–921. doi: 10.1210/en.2010-0981
Harber, M. P.,
Konopka, A. R., Douglass, M. D., Minchev, K., Kaminsky, L. A., Trappe, T. A.,
& Trappe, S. (2009). Aerobic exercise training improves whole muscle and
single myofiber size and function in older women. American Journal of
Physiology-Regulatory, Integrative and Comparative Physiology, 297(5). doi:
10.1152/ajpregu.00354.2009
Harber, M. P.,
Konopka, A. R., Undem, M. K., Hinkley, J. M., Minchev, K., Kaminsky, L. A., …
Trappe, S. (2012). Aerobic exercise training induces skeletal muscle
hypertrophy and age-dependent adaptations in myofiber function in young and
older men. Journal of Applied Physiology, 113(9), 1495–1504. doi:
10.1152/japplphysiol.00786.2012
Harridge, S.
(2007) Plasticity of human skeletal muscle: Gene expression to in vivo
function. Exp Physiolo. 92: 738-797.
Helms, E., Aragon,
A. & Fitschen, P. (2014). Evidence-based recommendations for natural
bodybuilding contest preparation: Nutrition and supplementation. Journal of the
International Society of Sports Nutrition. 11. 20. 10.1186/1550-2783-11-20.
Hill, M., Wernig,
A., & Goldspink, G. (2003). Muscle satellite (stem) cell activation during
local tissue injury and repair. Journal of Anatomy, 203(1), 89–99. doi:
10.1046/j.1469-7580.2003.00195.x
Hornberger, T. A.,
Chu, W. K., Mak, Y. W., Hsiung, J. W., Huang, S. A., & Chien, S. (2006).
The role of phospholipase D and phosphatidic acid in the mechanical activation
of mTOR signaling in skeletal muscle. Proceedings of the National Academy of Sciences,
103(12), 4741–4746. doi: 10.1073/pnas.0600678103
Izquierdo, M.,
Ibañez, J., González-Badillo, J. J., Häkkinen, K., Ratamess, N. A., Kraemer, W.
J., French, D., Eslava, J., Altadill, A., Asiain X. & Gorostiaga, E. M.
(2006). Differential effects of strength training leading to failure versus not
to failure on hormonal responses, strength, and muscle power gains. Journal of
Applied Physiology, 100(5), 1647–1656. doi: 10.1152/japplphysiol.01400.2005
Jacinto, E., &
Hall, M. N. (2003). TOR signalling in bugs, brain and brawn. Nature Reviews
Molecular Cell Biology, 4(2), 117–126. doi: 10.1038/nrm1018
Kadi, F.,
Eriksson, A., Holmner, S., Butler-Browne, G. & Thornell, L. (1999) Cellular
adaptation of the trapezius muscle in strength-trained athletes. Histochemistry
and cell biology, 111(3): 189-95.
Kadi, F. &
Thornell, L.E. (2000) Concomitant increases in myonuclear and satellite cell
content of female trapezius muscle following strength training. Histochemistry
and cell biology 113, 99-103.
Keogh, J. W.,
Wilson, G. J., & Weatherby, R. E. (1999). A Cross-Sectional Comparison of
Different Resistance Training Techniques in the Bench Press. Journal of
Strength and Conditioning Research, 13(3), 247–258. doi:
10.1519/00124278-199908000-00012
Kohn, T. A., Essén-Gustavsson,
B., & Myburgh, K. H. (2010). Specific muscle adaptations in type II fibers
after high-intensity interval training of well-trained runners. Scandinavian
Journal of Medicine & Science in Sports, 21(6), 765–772. doi:
10.1111/j.1600-0838.2010.01136.x
Kosek, D. J., Kim,
J.-S., Petrella, J. K., Cross, J. M., & Bamman, M. M. (2006). Efficacy of 3
days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in
young vs. older adults. Journal of Applied Physiology, 101(2), 531–544. doi:
10.1152/japplphysiol.01474.2005
Krieger, J. W.
(2010). Single vs. Multiple Sets of Resistance Exercise for Muscle Hypertrophy:
A Meta-Analysis. Journal of Strength and Conditioning Research, 24(4),
1150–1159. doi: 10.1519/jsc.0b013e3181d4d436
Lang, F (2007).
Mechanisms and Significance of Cell Volume Regulation. J Am Coll Nutr., 26:
613s-623s.
Lang, F., Busch,
G. L., Ritter, M., Völkl, H., Waldegger, S., Gulbins, E., & Häussinger, D.
(1998). Functional Significance of Cell Volume Regulatory Mechanisms.
Physiological Reviews, 78(1), 247–306. doi: 10.1152/physrev.1998.78.1.247
Loenneke, J. P.,
Wilson, J. M., Marín, P. J., Zourdos, M. C., & Bemben, M. G. (2011). Low
intensity blood flow restriction training: a meta-analysis. European Journal of
Applied Physiology, 112(5), 1849–1859. doi: 10.1007/s00421-011-2167-x
Loenneke, J. P.,
Thiebaud, R. S., & Abe, T. (2014). Does blood flow restriction result in
skeletal muscle damage? A critical review of available evidence. Scandinavian
Journal of Medicine & Science in Sports, 24(6). doi: 10.1111/sms.12210
Lynn, R. &
Morgan, D. (1994) Decline running produces more sarcomeres in rat vastus
intermedius muscle fibers than does incline running. Journal of applied
physiology: 77: 1439-1444.
Malm, C. (2001).
Exercise-induced muscle damage and inflammation: fact or fiction? Acta
Physiologica Scandinavica, 171(3), 233–239. doi:
10.1046/j.1365-201x.2001.00825.x
Macdougall, J. D.,
Gibala, M., Tarnopolsky, M., Macdonald, J., Interisano, S., & Yarasheski,
K. (1995). The Time Course For Elevated Muscle Protein Synthesis Following
Heavy Resistance Exercise. Medicine & Science in Sports & Exercise,
27(Supplement). doi: 10.1249/00005768-199505001-00367
MacDougall, D. Ward, G., Sale, D. & Sutton, J.
(1977) Biochemical adaptation of human skeletal muscle to heavy resistance
training and immobilization. Journal of applied physiology:
respiratory, environmental and exercise physiology: 43(4): 700-3.
Macdougall, J. D., Sale, D. G., Alway, S. E., &
Sutton, J. R. (1984). Muscle fiber number in biceps brachii in bodybuilders and
control subjects. Journal of Applied Physiology, 57(5), 1399–1403. doi:
10.1152/jappl.1984.57.5.1399
Mascher, H., Andersson, H., Nilsson, P.-A., Ekblom,
B., & Blomstrand, E. (2007). Changes in signalling pathways regulating
protein synthesis in human muscle in the recovery period after endurance
exercise. Acta Physiologica, 191(1), 67–75. doi: 10.1111/j.1748-1716.2007.01712.x
Masuda K., Choi J.Y., Shimojo H., Katsuta S. (1999) Maintenance of myoglobin concentration in human skeletal muscle
after heavy resistance training. European
Journal of Applied Physiology and Occupational Physiology 79(4), 347-352. [PubMed] [Google Scholar]
Martineau, L. & Gardiner,
P. (2002) Skeletal muscle is sensitive to the tension-time integral but not to
the rate of change of tension, as assessed by mechanically induced signaling.
Journal of biomechanic, 35: 657-663.
Mchugh, M. P. (2003). Recent advances in the
understanding of the repeated bout effect: the protective effect against muscle
damage from a single bout of eccentric exercise. Scandinavian Journal of
Medicine and Science in Sports, 13(2), 88–97. doi:
10.1034/j.1600-0838.2003.02477.x
Mcpherron, A. C., Lawler, A.
M., & Lee, S.-J. (1997). Regulation of skeletal muscle mass in mice by a
new TGF-p superfamily member. Nature, 387(6628), 83–90. doi:
10.1038/387083a0
Mikkola,
J., Rusko, H., Izquierdo, M., Gorostiaga, E., & Häkkinen, K. (2012). Neuromuscular and Cardiovascular Adaptations During
Concurrent Strength and Endurance Training in Untrained Men. International
Journal of Sports Medicine, 33(09), 702–710. doi: 10.1055/s-0031-1295475
Moore, D., Phillips, S., Babraj, J., Smith, K. & Rennie, M. (2005) Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. American journal of physiology - endocrinology andmetabolism, 288: E1153–E1159.
Monteiro, A., Aoki., M.,
Evangelista,A., Alveno, D., Monteiro, G., Picarro Ida, C. & Ugrinowitcch,
C. (2009) Nonlinear periodization maximizes strength gains in split resistance
training routines. Journal of strength and conditioning research, 23: 1321-1326.
Netreba, A., Popov, D., Bravyy,
Y., Lyubaeva, E., Terada, M., Ohira, T.,
Okabe, H., Vinogradova, O.& Ohira, Y. (2013). Responses of knee extensor
muscles to leg press training of various types in human. Rossiĭskii
fiziologicheskiĭ zhurnal imeni I.M. Sechenova / Rossiĭskaia akademiia nauk. 99.
406-16.
Nguyen, H. X., & Tidball,
J. G. (2003). Null Mutation of gp91phoxReduces Muscle Membrane Lysis During
Muscle Inflammation in Mice. The Journal of Physiology, 553(3), 833–841. doi:
10.1113/jphysiol.2003.051912
Nosaka, K., Lavender, A.,
Newton, M. & Sacco, P. (2003) Muscle damage in resistance training – is muscle
damage necessary for strength gain and muscle hypertrophy? International
journal of sport and health science, 1(1):1-8.
Nuckols, G. (2018)
Periodization: What the data say. https://www.strongerbyscience.com/periodization-data/
Ogasawara, R., Kobayashi, K.,
Tsutaki, A., Lee, K., Abe, T., Fujita, S., … Ishii, N. (2013). mTOR signaling
response to resistance exercise is altered by chronic resistance training and
detraining in skeletal muscle. Journal of Applied Physiology, 114(7), 934–940.
doi: 10.1152/japplphysiol.01161.2012
Pescatello, L. S., Devaney, J.
M., Hubal, M. J., Thompson, P. D., & Hoffman, E. P. (2013). Highlights from
the Functional Single Nucleotide Polymorphisms Associated with Human Muscle
Size and Strength or FAMuSS Study. BioMed Research International, 2013, 1–11.
doi: 10.1155/2013/643575
Petrella, J. K., Kim, J.-S.,
Mayhew, D. L., Cross, J. M., & Bamman, M. M. (2008). Potent myofiber
hypertrophy during resistance training in humans is associated with satellite
cell-mediated myonuclear addition: a cluster analysis. Journal of Applied
Physiology, 104(6), 1736–1742. doi: 10.1152/japplphysiol.01215.2007
Pinto, R., Gomes, N.,
Radaelli, R., Botton, C., Brown, L. & Bottaro, M. (2011). Effect of Range
of Motion on Muscle Strength and Thickness. Journal of strength and
conditioning research / National Strength & Conditioning Association. 26.
2140-5. 10.1519/JSC.0b013e31823a3b15.
Poliquin, C. (1988) Five steps
to increasing the effectiveness of your strength training program. Journal of
strength and conditioning association. 10: 34-39.
Prestes, J., Frollini, A. B.,
Lima, C. D., Donatto, F. F., Foschini, D., Marqueti, R. D. C., … Fleck, S. J.
(2009). Comparison Between Linear and Daily Undulating Periodized Resistance
Training to Increase Strength. Journal of Strength and Conditioning Research,
23(9), 2437–2442. doi: 10.1519/jsc.0b013e3181c03548
Qaisar, R., Bhaskaran, S.
& Remmen, H. (2016). Muscle
fiber type diversification during exercise and regeneration. Free Radical
Biology and Medicine. 98. 10.1016/j.freeradbiomed.2016.03.025.
Radaelli, R. J Fleck, S.,
Leite, T., Leite, R., Pinto, R., Fernandesm L. Simão, R. (2014). Dose Response
of 1, 3 and 5 Sets of Resistance Exercise on Strength, Local Muscular Endurance
and Hypertrophy. The Journal of Strength and Conditioning Research. 29.
10.1519/JSC.0000000000000758.
Ratamess, N. A., Falvo, M. J.,
Mangine, G. T., Hoffman, J. R., Faigenbaum, A. D., & Kang, J. (2007). The
effect of rest interval length on metabolic responses to the bench press
exercise. European Journal of Applied Physiology, 100(1), 1–17. doi:
10.1007/s00421-007-0394-y
Schoenfeld, B. (2010) The
mechanism of muscle hypertrophy and their application to resistance training.
Journal of strength and conditioning research, 24: 2857-2872.
Schoenfeld, BJ. Science and Development of Muscle Hypertrophy. Champaign, IL; Human Kinetics, 2016.
Schoenfeld, B, Ogborn, D.
& Krieger, J. (2017) Dose-response relationship between weekly resistance
training volume and increases in muscle mass: A systematic review and
meta-analysis, Journal of Sports Sciences, 35:11, 1073-1082, DOI:
10.1080/02640414.2016.1210197
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., Sonmez, G. T., & Alvar, B. A.
(2014). Effects of Different Volume-Equated Resistance Training Loading
Strategies on Muscular Adaptations in Well-Trained Men. Journal of Strength and
Conditioning Research, 28(10), 2909–2918. doi: 10.1519/jsc.0000000000000480
Schoenfeld, B, Grgic, J.,
Ogbord, D. & Krieger, J. (2017) Strength and hypertrophy adaptations
between low- versus high-load resistance training: A systematic review and
meta-analysis. The Journal of strength and conditioning research, Ahead of
Print DOI: 10.1519/JSC.0000000000002200
Schoenfeld, B., Contreras, B.,
Vigotsky, A., Ogborn, D., Fontana, F. & Tiryaki-Sonmez, R. (2016). Upper
body muscle activation during low-versus high-load resistance exercise in the
bench press. 24. 217-224. 10.3233/IES-160620.
Schoenfeld, B. J., Ratamess,
N. A., Peterson, M. D., Contreras, B., & Tiryaki-Sonmez, G. (2015).
Influence of Resistance Training Frequency on Muscular Adaptations in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(7),
1821–1829. doi: 10.1519/jsc.0000000000000970
Schoenfeld, B. J., Peterson,
M. D., Ogborn, D., Contreras, B., & Sonmez, G. T. (2015). Effects of Low-
vs. High-Load Resistance Training on Muscle Strength and Hypertrophy in
Well-Trained Men. Journal of Strength and Conditioning Research, 29(10), 2954–2963.
doi: 10.1519/jsc.0000000000000958
Schoenfeld, B. J., Ogborn, D.
I., & Krieger, J. W. (2015). Effect of Repetition Duration During
Resistance Training on Muscle Hypertrophy: A Systematic Review and
Meta-Analysis. Sports Medicine, 45(4), 577–585. doi: 10.1007/s40279-015-0304-0
Schoenfeld, B. J., Contreras,
B., Krieger, J., Grgic, J., Delcastillo, K., Belliard, R., & Alto, A.
(2019). Resistance Training Volume Enhances Muscle Hypertrophy but Not Strength
in Trained Men. Medicine & Science in Sports & Exercise, 51(1), 94–103.
doi: 10.1249/mss.0000000000001764
Schoenfeld, B.J. & Grgic,
J. (2020) Effects of range of motion on muscle development during resistance
training interventions: A systematic review. SAGE open medicine, 8: 1-8. https://doi.org/10.1177/2050312120901559
Schuenke, M. D., Herman, J.
R., Gliders, R. M., Hagerman, F. C., Hikida, R. S., Rana, S. R., … Staron, R.
S. (2012). Early-phase muscular adaptations in response to slow-speed versus
traditional resistance-training regimens. European Journal of Applied
Physiology, 112(10), 3585–3595. doi: 10.1007/s00421-012-2339-3
Selye, H. (1950). Stress and
the General Adaptation Syndrome. Bmj, 1(4667), 1383–1392. doi:
10.1136/bmj.1.4667.1383
Serrano, A. L., Baeza-Raja,
B., Perdiguero, E., Jardí, M., & Muñoz-Cánoves, P. (2008). Interleukin-6 Is
an Essential Regulator of Satellite Cell-Mediated Skeletal Muscle Hypertrophy.
Cell Metabolism, 7(1), 33–44. doi: 10.1016/j.cmet.2007.11.011
Simão, R., Spineti, J.,
Salles, B. F. D., Matta, T., Fernandes, L., Fleck, S. J., … Strom-Olsen, H. E.
(2012). Comparison Between Nonlinear and Linear Periodized Resistance Training.
Journal of Strength and Conditioning Research, 26(5), 1389–1395. doi:
10.1519/jsc.0b013e318231a659
Snyder, B. J., & Leech, J.
R. (2009). Voluntary Increase in Latissimus Dorsi Muscle Activity During the
Lat Pull-Down Following Expert Instruction. Journal of Strength and
Conditioning Research, 23(8), 2204–2209. doi: 10.1519/jsc.0b013e3181bb7213
Snyder, B. J., & Fry, W.
R. (2012). Effect of Verbal Instruction on Muscle Activity During the Bench
Press Exercise. Journal of Strength and Conditioning Research, 26(9), 2394–2400.
doi: 10.1519/jsc.0b013e31823f8d11
Stewart, C. & Rittweger,
J. (2006) Adaptive processes in skeletal muscle: molecular regulators and
genetic influences. Journal of musculoskeletal and neuronal interactions: 6(1): 73-86.
Stone, M., O’Bryant, H. &
Garhammer J. (1981) Hypothetical model for strength training. Journal of sports
medicine and physical fitness, 21: 342-351.
Stone, M. H., Potteiger, J.
A., Pierce, K. C., Proulx, C. M., Obryant, H. S., Johnson, R. L., & Stone,
M. E. (2000). Comparison of the Effects of Three Different Weight-Training
Programs on the One Repetition Maximum Squat. The Journal of Strength and
Conditioning Research, 14(3), 332. doi:
10.1519/1533-4287(2000)014<0332:coteot>2.0.co;2
Takarada,
Y., Takazawa, H., Sato, Y., Takebayashi, S., Tanaka, Y., & Ishii, N.
(2000). Effects of
resistance exercise combined with moderate vascular occlusion on muscular
function in humans. Journal of Applied Physiology, 88(6), 2097–2106. doi:
10.1152/jappl.2000.88.6.2097
Takarada, Y., Nakamura, Y.,
Aruga, S., Onda, T., Miyazaki, S., & Ishii, N. (2000). Rapid increase in plasma growth hormone after
low-intensity resistance exercise with vascular occlusion. Journal of Applied
Physiology, 88(1), 61–65. doi: 10.1152/jappl.2000.88.1.61
Tee, J. C., Bosch, A. N.,
& Lambert, M. I. (2007). Metabolic Consequences of Exercise-Induced Muscle
Damage. Sports Medicine, 37(10), 827–836. doi: 10.2165/00007256-200737100-00001
Terzis, G., Spengos, K., Mascher,
H., Georgiadis, G., Manta, P. &
Blomstrand, E. (2010). The degree of p70S6k and S6 phosphorylation in human
skeletal muscle in response to resistance exercise depends on the training
volume. European journal of applied physiology. 110. 835-43.
10.1007/s00421-010-1527-2.
Thomas, G., & Hall, M. N.
(1997). TOR signalling and control of cell growth. Current Opinion in Cell
Biology, 9(6), 782–787. doi: 10.1016/s0955-0674(97)80078-6
Uchiyama, S., Tsukamoto, H.,
Yoshimura, S., & Tamaki, T. (2006). Relationship between oxidative stress in muscle tissue
and weight-lifting-induced muscle damage. Pflügers Archiv - European Journal of
Physiology, 452(1), 109–116. doi: 10.1007/s00424-005-0012-y
Wernbom, M., Augustsson, J.
& Thomee, R. (2007). The Influence of Frequency, Intensity, Volume and Mode
of Strength Training on Whole Muscle Cross-Sectional Area in Humans. Sports
medicine (Auckland, N.Z.). 37. 225-64. 10.2165/00007256-200737030-00004.
Willoughby, D. (1993) The
effects of mesocycle-length weight training programs involving periodization an
partially equated volumes on upper and lower body strength. Journal of strength
and conditioning research, 7: 2-8.
Wilson, J. M., Lowery, R. P.,
Joy, J. M., Loenneke, J. P., & Naimo, M. A. (2013). Practical Blood Flow
Restriction Training Increases Acute Determinants of Hypertrophy Without
Increasing Indices of Muscle Damage. Journal of Strength and Conditioning
Research, 27(11), 3068–3075. doi: 10.1519/jsc.0b013e31828a1ffa
Wilson, J. M., Marin, P. J.,
Rhea, M. R., Wilson, S. M., Loenneke, J. P., & Anderson, J. C. (2012).
Concurrent Training. Journal of Strength and Conditioning Research, 26(8),
2293–2307. doi: 10.1519/jsc.0b013e31823a3e2d
Wulf, G. (2013). Attentional
focus and motor learning: a review of 15 years. International Review of Sport
and Exercise Psychology, 6(1), 77–104. doi: 10.1080/1750984x.2012.723728
Yang, S. Y., & Goldspink,
G. (2002). Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in
myoblast proliferation and differentiation. FEBS Letters, 522(1-3), 156–160.
doi: 10.1016/s0014-5793(02)02918-6
Zanou, N., & Gailly, P.
(2013). Skeletal muscle hypertrophy and regeneration: interplay between the
myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs)
pathways. Cellular and Molecular Life Sciences, 70(21), 4117–4130. doi:
10.1007/s00018-013-1330-4
Zanchi, N. E., & Lancha,
A. H. (2007). Mechanical stimuli of skeletal muscle: implications on
mTOR/p70s6k and protein synthesis. European Journal of Applied Physiology,
102(3), 253–263. doi: 10.1007/s00421-007-0588-3
Olen pääkaupunkiseudulla vaikuttava voima-, fysiikka- ja yleisurheiluvalmentaja. Valmentajana olen toiminut kohta 10 vuoden ajan. Olen erityisen kiinnostunut urheilijoiden suorituskyvystä ja harjoittelun yksilöllisistä eroista urheilijoilla sekä periodisaatiosta. Saan urheilijat suoriutumaan lajissaan paremmin.
Tämä sivusto on luotu aputyökaluksi valmentajille, jotka haluavat perustaa valmentamisensa tieteelliseen näyttöön. Alalla on vielä paljon käytössä vanhentuneita toimintatapoja, jotka heikentävät urheilijoiden suorituskykyä. Pyrin jakamaan tieteellistä tietoa ja auttamaan objektiivisesti kaikkia valmentajia.